2005 Book Mountainecosystems.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Chapter14.Pdf
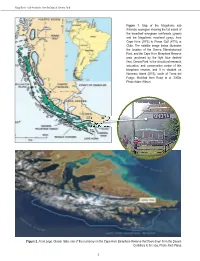
PART I • Omora Park Long-Term Ornithological Research Program THE OMORA PARK LONG-TERM ORNITHOLOGICAL RESEARCH PROGRAM: 1 STUDY SITES AND METHODS RICARDO ROZZI, JAIME E. JIMÉNEZ, FRANCISCA MASSARDO, JUAN CARLOS TORRES-MURA, AND RAJAN RIJAL In January 2000, we initiated a Long-term Ornithological Research Program at Omora Ethnobotanical Park in the world's southernmost forests: the sub-Antarctic forests of the Cape Horn Biosphere Reserve. In this chapter, we first present some key climatic, geographical, and ecological attributes of the Magellanic sub-Antarctic ecoregion compared to subpolar regions of the Northern Hemisphere. We then describe the study sites at Omora Park and other locations on Navarino Island and in the Cape Horn Biosphere Reserve. Finally, we describe the methods, including censuses, and present data for each of the bird species caught in mist nets during the first eleven years (January 2000 to December 2010) of the Omora Park Long-Term Ornithological Research Program. THE MAGELLANIC SUB-ANTARCTIC ECOREGION The contrast between the southwestern end of South America and the subpolar zone of the Northern Hemisphere allows us to more clearly distinguish and appreciate the peculiarities of an ecoregion that until recently remained invisible to the world of science and also for the political administration of Chile. So much so, that this austral region lacked a proper name, and it was generally subsumed under the generic name of Patagonia. For this reason, to distinguish it from Patagonia and from sub-Arctic regions, in the early 2000s we coined the name “Magellanic sub-Antarctic ecoregion” (Rozzi 2002). The Magellanic sub-Antarctic ecoregion extends along the southwestern margin of South America between the Gulf of Penas (47ºS) and Horn Island (56ºS) (Figure 1). -

Bioclimatic and Phytosociological Diagnosis of the Species of the Nothofagus Genus (Nothofagaceae) in South America
International Journal of Geobotanical Research, Vol. nº 1, December 2011, pp. 1-20 Bioclimatic and phytosociological diagnosis of the species of the Nothofagus genus (Nothofagaceae) in South America Javier AMIGO(1) & Manuel A. RODRÍGUEZ-GUITIÁN(2) (1) Laboratorio de Botánica, Facultad de Farmacia, Universidad de Santiago de Compostela (USC). E-15782 Santiago de Com- postela (Galicia, España). Phone: 34-881 814977. E-mail: [email protected] (2) Departamento de Producción Vexetal. Escola Politécnica Superior de Lugo-USC. 27002-Lugo (Galicia, España). E-mail: [email protected] Abstract The Nothofagus genus comprises 10 species recorded in the South American subcontinent. All are important tree species in the ex- tratropical, Mediterranean, temperate and boreal forests of Chile and Argentina. This paper presents a summary of data on the phyto- coenotical behaviour of these species and relates the plant communities to the measurable or inferable thermoclimatic and ombrocli- matic conditions which affect them. Our aim is to update the phytosociological knowledge of the South American temperate forests and to assess their suitability as climatic bioindicators by analysing the behaviour of those species belonging to their most represen- tative genus. Keywords: Argentina, boreal forests, Chile, mediterranean forests, temperate forests. Introduction tually give rise to a temperate territory with rainfall rates as high as those of regions with a Tropical pluvial bio- The South American subcontinent is usually associa- climate; iii. finally, towards the apex of the American ted with a tropical environment because this is in fact the Southern Cone, this temperate territory progressively dominant bioclimatic profile from Panamá to the north of gives way to a strip of land with a Boreal bioclimate. -

Temporal Change in Tree Species Composition in Palampur Forest
2019 Status Report Palampur Palampur Forest Division Temporal Change in Tree Species Composition in Palampur Forest Division of Dharamshala Forest Circle, Himachal Pradesh Harish Bharti, Aditi Panatu, Kiran and Dr. S. S. Randhawa H. P. State Centre on Climate Change (HIMCOSTE), Vigyan Bhawan near Udyog Bhawan, Bemloe Shimla-01 0177-2656489 Table of Contents Introduction ............................................................................................................................................ 4 Forests of Himachal Pradesh........................................................................................................................ 5 Study area and method ....................................................................................................................... 7 District Kangra A Background .................................................................................................................. 7 Location & Geographical– Area ................................................................................................................. 8 Palampur Forest Division- Forest Profile................................................................................................ 9 Name and Situation:- .................................................................................................................................. 9 Geology: ......................................................................................................................................................... 11 -

The Role of Leaf Cellulose Content in Determining Host Plant Preferences of Three Defoliating Insects Present in the Andean-Patagonian Forest
Austral Ecology (2016) , – The role of leaf cellulose content in determining host plant preferences of three defoliating insects present in the Andean-Patagonian forest A. L. PIETRANTUONO, O. A. BRUZZONE AND V. FERNANDEZ-ARHEX* Estacion Experimental Agropecuaria Bariloche, CONICET – Instituto Nacional de Tecnologıa Agropecuaria, CC277, Av. Modesta Victoria 4450, 8400 San Carlos de Bariloche, Rıo Negro, Argentina (E-mail: [email protected]) Abstract Phytophagous insects choose their feeding resources according to their own requirements in addition to properties of the host plants, such as biomechanical defences. The feeding preferences of the native folivorous insects of the Andean-Patagonian forest (Argentina) have rarely been studied. These environments present a wide diversity and abundance of insects associated with trees of the Nothofagus and Lophozonia (Nothofagaceae) genera, which represent the main tree species of the forests of the southern hemisphere. In particular, Lophozonia alpina and Lophozonia obliqua are of great interest because they have a wide distribution, a high capacity for hybridization and exhibit great phenotypic plasticity. This versatility causes substantial variation in the biome- chanical properties of leaves, affecting the feeding preferences of insects. The purpose of this work was to study the food selection behaviour of three leaf-chewing insects (Polydrusus nothofagii, Polydrusus roseaus (Coleoptera: Curculionidae) and Perzelia arda (Lepidoptera: Oecophoridae)) associated with L. alpina and L. obliqua as host plants. Based on their choices, our aim was to determine a preference scale for each insect species and the vari- ables on which these preferences were based. Therefore, we selected trees of L. alpina and L. obliqua, measured several properties such as cellulose content and recorded which leaves were eaten. -

Principles and Practice of Forest Landscape Restoration Case Studies from the Drylands of Latin America Edited by A.C
Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor About IUCN IUCN, International Union for Conservation of Nature, helps the world find pragmatic solutions to our most pressing environment and development challenges. IUCN works on biodiversity, climate change, energy, human livelihoods and greening the world economy by supporting scientific research, managing field projects all over the world, and bringing governments, NGOs, the UN and companies together to develop policy, laws and best practice. IUCN is the world’s oldest and largest global environmental organization, with more than 1,000 government and NGO members and almost 11,000 volunteer experts in some 160 countries. IUCN’s work is supported by over 1,000 staff in 60 offices and hundreds of partners in public, NGO and private sectors around the world. www.iucn.org Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor This book is dedicated to the memory of Margarito Sánchez Carrada, a student who worked on the research project described in these pages. The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN or the European Commission concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Bioclimatic and Phytosociological Diagnosis of the Species of the Nothofagus Genus (Nothofagaceae) in South America
International Journal of Geobotanical Research, Vol. nº 1, December 2011, pp. 1-20 Bioclimatic and phytosociological diagnosis of the species of the Nothofagus genus (Nothofagaceae) in South America Javier AMIGO(1) & Manuel A. RODRÍGUEZ-GUITIÁN(2) (1) Laboratorio de Botánica, Facultad de Farmacia, Universidad de Santiago de Compostela (USC). E-15782 Santiago de Com- postela (Galicia, España). Phone: 34-881 814977. E-mail: [email protected] (2) Departamento de Producción Vexetal. Escola Politécnica Superior de Lugo-USC. 27002-Lugo (Galicia, España). E-mail: [email protected] Abstract The Nothofagus genus comprises 10 species recorded in the South American subcontinent. All are important tree species in the ex- tratropical, Mediterranean, temperate and boreal forests of Chile and Argentina. This paper presents a summary of data on the phyto- coenotical behaviour of these species and relates the plant communities to the measurable or inferable thermoclimatic and ombrocli- matic conditions which affect them. Our aim is to update the phytosociological knowledge of the South American temperate forests and to assess their suitability as climatic bioindicators by analysing the behaviour of those species belonging to their most represen- tative genus. Keywords: Argentina, boreal forests, Chile, mediterranean forests, temperate forests. Introduction tually give rise to a temperate territory with rainfall rates as high as those of regions with a Tropical pluvial bio- The South American subcontinent is usually associa- climate; iii. finally, towards the apex of the American ted with a tropical environment because this is in fact the Southern Cone, this temperate territory progressively dominant bioclimatic profile from Panamá to the north of gives way to a strip of land with a Boreal bioclimate. -

Principles and Practice of Forest Landscape Restoration Case Studies from the Drylands of Latin America Edited by A.C
Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor About IUCN IUCN, International Union for Conservation of Nature, helps the world find pragmatic solutions to our most pressing environment and development challenges. IUCN works on biodiversity, climate change, energy, human livelihoods and greening the world economy by supporting scientific research, managing field projects all over the world, and bringing governments, NGOs, the UN and companies together to develop policy, laws and best practice. IUCN is the world’s oldest and largest global environmental organization, with more than 1,000 government and NGO members and almost 11,000 volunteer experts in some 160 countries. IUCN’s work is supported by over 1,000 staff in 60 offices and hundreds of partners in public, NGO and private sectors around the world. www.iucn.org Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Principles and Practice of Forest Landscape Restoration Case studies from the drylands of Latin America Edited by A.C. Newton and N. Tejedor This book is dedicated to the memory of Margarito Sánchez Carrada, a student who worked on the research project described in these pages. The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN or the European Commission concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Burlington House
Sutainable Resource Development in the Himalaya Contents Pages 2-5 Oral Programme Pages 6-7 Poster programme Pages 8-33 Oral presentation abstracts (in programme order) Pages 34-63 Poster presentations abstracts (in programme order) Pages 64-65 Conference sponsor information Pages 65-68 Notes 24-26 June 2014 Page 1 Sutainable Resource Development in the Himalaya Oral Programme Tuesday 24 June 2014 09.00 Welcome 11.30 Student presentation from Leh School 11.45 A life in Ladakh Professor (ambassador) Phunchok Stobdan, Institute for Defence Studies and Analyses 12.30 Lunch and posters 14.00 Mountaineering in the Himalaya Ang Rita Sherpa, Mountain Institute, Kathmandu, Nepal Session theme: The geological framework of the Himalaya 14.30 Geochemical and isotopic constraints on magmatic rocks – some constraints on collision based on new SHRIMP data Professor Talat Ahmed, University of Kashmir 15.15 Short subject presentations and panel discussion Moderators: Director, Geology & Mining, Jammu & Kashmir State & Director, Geological Survey of India Structural framework of the Himalayas with emphasis on balanced cross sections Professor Dilip Mukhopadhyay, IIT Roorkee Sedimentology Professor S. K. Tandon, Delhi University Petrogenesis and economic potential of the Early Permian Panjal Traps, Kashmir, India Mr Greg Shellnut, National Taiwan Normal University Precambrian Professor D. M. Banerjee, Delhi University 16.00 Tea and posters 16.40 Short subject presentations continued & panel discussion 18.00 Close of day 24-26 June 2014 Page 2 Sutainable Resource Development in the Himalaya Wednesday 25 June 2014 Session theme: Climate, Landscape Evolution & Environment 09.00 Climate Professor Harjeet Singh, JNU, New Delhi 09.30 Earth surface processes and landscape evolution in the Himalaya Professor Lewis Owen, Cincinnati University 10.00 Landscape & Vegetation Dr P. -

Argentine and Chilean Claims to British Antarctica. - Bases Established in the South Shetlands
Keesing's Record of World Events (formerly Keesing's Contemporary Archives), Volume VI-VII, February, 1948 Argentine, Chilean, British, Page 9133 © 1931-2006 Keesing's Worldwide, LLC - All Rights Reserved. Argentine and Chilean Claims to British Antarctica. - Bases established in the South Shetlands. - Chilean President inaugurates Chilean Army Bases on Greenwich Island. - Argentine Naval Demonstration in British Antarctic Waters. - H.M.S. "Nigeria" despatched to Falklands. - British Government Statements. - Argentine-Chilean Agreement on Joint Defence of "Antarctic Rights." - The Byrd and Ronne Antarctic Expeditions. - Australian Antarctic Expedition occupies Heard Islands. The Foreign-Office in London, in statements on Feb. 7 and Feb. 13, announced that Argentina and Chile had rejected British protests, earlier presented in Buenos Aires and Santiago, against the action of those countries in establishing bases in British Antarctic territories. The announcement of Feb. 7 stated that on Dec. 7, 1947, the British Ambassador in Buenos Aires, Sir Reginald Leeper, had presented a Note expressing British "anxiety" at the activities in the Antarctic of an Argentine naval expedition which had visited part of the Falkland Islands Dependencies, including Graham Land, the South Shetlands, and the South Orkneys, and had landed at various points in British territory; that a request had been made for Argentine nationals to evacuate bases established on Deception Island and Gamma Island, in the South Shetlands; that H.M. Government had proposed that the Argentine should submit her claim to Antarctic sovereignty to the International Court of Justice for adjudication; and that on Dec. 23, 1947, a second British Note had been presented expressing surprise at continued violations of British territory and territorial waters by Argentine vessels in the Antarctic. -

Pakistan K7 Attempt. Japanese Led by Masayuki Hoshina Made an Attempt on K7 (6934 Meters, 22,750 Feet) by Way of the 17,000-Foot West Col
268 THE AMERICAN KPINE JOURNAL. I983 Pakistan K7 Attempt. Japanese led by Masayuki Hoshina made an attempt on K7 (6934 meters, 22,750 feet) by way of the 17,000-foot west col. They ap- proached from Hushe via the Charakusa Glacier, where they established Base Camp on May 27. They fixed some 5000 feet of rope. The expedition reached a little higher than 20,000 feet. Hidden Peak (Gasherbrum I), North Face Attempt. Granger Banks, Rich- ard Soaper, Lyle Dean and I arrived at Gasherbrum I Base Camp on May 19 after eleven days on the Baltoro approach with 23 porters. After placing a food cache at 21,325 feet in the Gasherbrum La Icefall, we descendedto recuperate. On June 3 we returned up the West Gasherbrum Glacier icefall to the cache in two days. The next day we climbed the right side of the north face in twelve hours unroped. The face consisted of wind-blown ice on the bottom, mixed climbing on a rotten rock a&e in the middle and a final third of “funky” n&C up to 80”. We carried four days’ food. We were on the north face itself, well left of Messner’s route. On the top of the north face, on the northwest shoulder of Hidden Peak, we placed our high camp at 23,300 feet next to Messner’s and Habeler’s destroyed tent. On June 6 we rested, hoping to join Messner’s route the next day and climb to the summit. We were wrong. The next three days were spent battling gale winds. -

Kinematics of the Karakoram-Kohistan Suture Zone, Chitral, NW Pakistan
Research Collection Doctoral Thesis Kinematics of the Karakoram-Kohistan Suture Zone, Chitral, NW Pakistan Author(s): Heuberger, Stefan Publication Date: 2004 Permanent Link: https://doi.org/10.3929/ethz-a-004906874 Rights / License: In Copyright - Non-Commercial Use Permitted This page was generated automatically upon download from the ETH Zurich Research Collection. For more information please consult the Terms of use. ETH Library DISS. ETH NO. 15778 KINEMATICS OF THE KARAKORAM-KOHISTAN SUTURE ZONE, CHITRAL, NW PAKISTAN A dissertation submitted to the SWISS FEDERAL INSTITUTE OF TECHNOLOGY ZURICH for the degree of Doctor of Natural Sciences presented by STEFAN HEUBERGER Dipl. Natw. ETH Zürich born on August 6, 1976 citizen of Sirnach (TG), Rickenbach (TG) and Wilen (TG) accepted on the recommendation of Prof. Dr. J.-P. Burg ETH Zürich examiner Prof. Dr. U. Schaltegger Université de Genève co-examiner Prof. Dr. A. Zanchi Università di Milano co-examiner 2004 “Die verstehen sehr wenig, die nur das verstehen, was sich erklären lässt” Marie von Ebner-Eschenbach Acknowledgements Thanks: Daniel Bernoulli, Universität Basel; Jean-Louis Bodinier, ISTEEM Montpellier (F); Martin Bruderer, ETH Zürich; Jean-Pierre Burg, ETH Zürich; Bernard Célérier, ISTEEM Montpellier (F); Nawaz Muhammad Chaudhry, University of the Punjab, Lahore (PK); Nadeem’s cousin, Mansehra (PK); Hamid Dawood, PMNH Islamabad (PK); Mohammed Dawood, Madaglasht (PK); Yamina Elmer, St.Gallen; Martin Frank, ETH Zürich; Maurizio Gaetani, Università degli Studi di Milano (I); the family -

Invaders Without Frontiers: Cross-Border Invasions of Exotic Mammals
Biological Invasions 4: 157–173, 2002. © 2002 Kluwer Academic Publishers. Printed in the Netherlands. Review Invaders without frontiers: cross-border invasions of exotic mammals Fabian M. Jaksic1,∗, J. Agust´ın Iriarte2, Jaime E. Jimenez´ 3 & David R. Mart´ınez4 1Center for Advanced Studies in Ecology & Biodiversity, Pontificia Universidad Catolica´ de Chile, Casilla 114-D, Santiago, Chile; 2Servicio Agr´ıcola y Ganadero, Av. Bulnes 140, Santiago, Chile; 3Laboratorio de Ecolog´ıa, Universidad de Los Lagos, Casilla 933, Osorno, Chile; 4Centro de Estudios Forestales y Ambientales, Universidad de Los Lagos, Casilla 933, Osorno, Chile; ∗Author for correspondence (e-mail: [email protected]; fax: +56-2-6862615) Received 31 August 2001; accepted in revised form 25 March 2002 Key words: American beaver, American mink, Argentina, Chile, European hare, European rabbit, exotic mammals, grey fox, muskrat, Patagonia, red deer, South America, wild boar Abstract We address cross-border mammal invasions between Chilean and Argentine Patagonia, providing a detailed history of the introductions, subsequent spread (and spread rate when documented), and current limits of mammal invasions. The eight species involved are the following: European hare (Lepus europaeus), European rabbit (Oryctolagus cuniculus), wild boar (Sus scrofa), and red deer (Cervus elaphus) were all introduced from Europe (Austria, France, Germany, and Spain) to either or both Chilean and Argentine Patagonia. American beaver (Castor canadensis) and muskrat (Ondatra zibethicus) were introduced from Canada to Argentine Tierra del Fuego Island (shared with Chile). The American mink (Mustela vison) apparently was brought from the United States of America to both Chilean and Argentine Patagonia, independently. The native grey fox (Pseudalopex griseus) was introduced from Chilean to Argentine Tierra del Fuego.