Recent Advances in Understanding the Mechanisms of Cerebellar
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ultrastructural Study of the Granule Cell Domain of the Cochlear Nucleus in Rats: Mossy Fiber Endings and Their Targets
THE JOURNAL OF COMPARATIVE NEUROLOGY 369~345-360 ( 1996) Ultrastructural Study of the Granule Cell Domain of the Cochlear Nucleus in Rats: Mossy Fiber Endings and Their Targets DIANA L. WEEDMAN, TAN PONGSTAPORN, AND DAVID K. RYUGO Center for Hearing Sciences, Departments of Otolaryngoloby-Head and Neck Surgery and Neuroscience, Johns Hopkins University School of Medicine, Baltimore, Maryland 2 1205 ABSTRACT The principal projection neurons of the cochlear nucleus receive the bulk of their input from the auditory nerve. These projection neurons reside in the core of the nucleus and are surrounded by an external shell, which is called the granule cell domain. Interneurons of the cochlear granule cell domain are the target for nonprimary auditory inputs, including projections from the superior olivary complex, inferior colliculus, and auditory cortex. The granule cell domain also receives projections from the cuneate and trigeminal nuclei, which are first-order nuclei of the somatosensory system. The cellular targets of the nonprimary projections are mostly unknown due to a lack of information regarding postsynaptic profiles in the granule cell areas. In the present paper, we examined the synaptic relationships between a heterogeneous class of large synaptic terminals called mossy fibers and their targets within subdivisions of the granule cell domain known as the lamina and superficial layer. By using light and electron microscopic methods in these subdivisions, we provide evidence for three different neuron classes that receive input from the mossy fibers: granule cells, unipolar brush cells, and a previously undescribed class called chestnut cells. The distinct synaptic relations between mossy fibers and members of each neuron class further imply fundamentally separate roles for processing acoustic signals. -

Consensus Paper: Cerebellar Development
Cerebellum DOI 10.1007/s12311-015-0724-2 CONSENSUS PAPER Consensus Paper: Cerebellar Development Ketty Leto1,2 & Marife Arancillo3 & Esther B. E. Becker4 & Annalisa Buffo1,2 & Chin Chiang5 & Baojin Ding6 & William B. Dobyns 7,8 & Isabelle Dusart9,10 & Parthiv Haldipur7 & Mary E. Hatten11 & Mikio Hoshino12 & Alexandra L. Joyner13 & Masanobu Kano14 & Daniel L. Kilpatrick6 & Noriyuki Koibuchi15 & Silvia Marino16 & Salvador Martinez17 & Kathleen J. Millen7 & Thomas O. Millner16 & Takaki Miyata18 & Elena Parmigiani1,2 & Karl Schilling19 & Gabriella Sekerková20 & Roy V. Sillitoe3 & Constantino Sotelo21 & Naofumi Uesaka14 & Annika Wefers 22 & Richard J. T. Wingate23 & Richard Hawkes24 # The Author(s) 2015. This article is published with open access at Springerlink.com Abstract The development of the mammalian cerebellum is processes of cerebellar ontogenesis, highlighting the neuro- orchestrated by both cell-autonomous programs and inductive genic strategies used by developing progenitors, the genetic environmental influences. Here, we describe the main programs involved in cell fate specification, the progressive * Ketty Leto 10 Centre National de la Recherche Scientifique, CNRS, UMR8246, [email protected] INSERM U1130, Neuroscience Paris Seine, France, 75005 Paris, France 11 Laboratory of Developmental Neurobiology, The Rockefeller 1 Department of Neuroscience Rita Levi Montalcini, University of University, New York, NY 10065, USA Turin, via Cherasco 15, 10026 Turin, Italy 12 Department of Biochemistry and Cellular Biology, National Institute -

Purkinje Cell Migration Disorder By
CEREBELLAR CORTICOGENESIS IN THE LYSOSOMAL ACID PHOSPHATASE (ACP2) MUTANT MICE: PURKINJE CELL MIGRATION DISORDER BY NILOUFAR ASHTARI A Thesis Submitted to the Faculty of Graduate Studies of The University of Manitoba in Partial Fulfilment of the Requirements for the Degree of MASTER OF SCIENCE Department of Human Anatomy and Cell Science University of Manitoba Winnipeg, Manitoba Copyright © 2017 by Niloufar Ashtari 1 Abstract In a mutant mouse called nax as the result of mutation in Lysosomal Acid phosphatase (Acp2), layers of the cerebellar cortex are impaired and monolayer Purkinje cells (Pcs) turn to multi-layered Pcs that ectopically invade the molecular layer. We investigated reelin-Dab1 signaling as an important pathway for Pcs migration and monolayer formation in cerebellum. ERK1/2 is a member of mitogen activated kinases family and suggested to be a downstream of reelin signaling. We hypothesize that the establishment of mono-layered Pcs rely on reelin through ERK1/2 pathway. Acp2 mutant mice were used for this study and molecular expression and distribution were assessed by immunohistochemistry, RT-PCR, western blotting, and cell culture. Results suggest that reelin may modulate the ERK1/2 expression, thus lower expression of reelin and higher phosphorylation of Dab1 leads to over expression of the ERK1/2 that causes the Pcs to over migrate and form multilayer in nax cerebellar cortex. i TABLE OF CONTENTS LISTOFABBREVIATIONS………………………………………………..…... IV LIST OF TABLES……………………………………...…………………...….. Vii LIST OF FIGURES…………………………………………………….………. Viii CHAPTER 1: INTRODUCTION…………………………………….………… 1 1.1 Cerebellum ……………………………………………………........………. 1 1.2 Development of Central Nervous System………………………………….. 2 1.3 Development of the cerebellum………………………………….................. 3 1.4 Specification of cerebellar germinal zones…………………………………. -

Parameters of Starch Granule Genesis in Chloroplasts of Arabidopsis Thaliana
Mathematisch-Naturwissenschaftliche Fakultät Irina Malinova | Hadeel M. Qasim | Henrike Brust | Joerg Fettke Parameters of Starch Granule Genesis in Chloroplasts of Arabidopsis thaliana Suggested citation referring to the original publication: Frontiers in Plant Science 9 (2018) Art, 761 DOI http://dx.doi.org/10.3389/fpls.2018.00761 ISSN (online) 1664-462X Postprint archived at the Institutional Repository of the Potsdam University in: Postprints der Universität Potsdam Mathematisch-Naturwissenschaftliche Reihe ; 478 ISSN 1866-8372 http://nbn-resolving.de/urn:nbn:de:kobv:517-opus4-419295 fpls-09-00761 June 3, 2018 Time: 11:48 # 1 MINI REVIEW published: 05 June 2018 doi: 10.3389/fpls.2018.00761 Parameters of Starch Granule Genesis in Chloroplasts of Arabidopsis thaliana Irina Malinova†, Hadeel M. Qasim, Henrike Brust† and Joerg Fettke* Biopolymer Analytics, University of Potsdam, Potsdam, Germany Starch is the primary storage carbohydrate in most photosynthetic organisms and allows the accumulation of carbon and energy in form of an insoluble and semi-crystalline particle. In the last decades large progress, especially in the model plant Arabidopsis thaliana, was made in understanding the structure and metabolism of starch and its conjunction. The process underlying the initiation of starch granules remains obscure, Edited by: although this is a fundamental process and seems to be strongly regulated, as in Yasunori Nakamura, Akita Prefectural University, Japan Arabidopsis leaves the starch granule number per chloroplast is fixed with 5-7. Several Reviewed by: single, double, and triple mutants were reported in the last years that showed massively Christophe D’Hulst, alterations in the starch granule number per chloroplast and allowed further insights in Lille University of Science and Technology, France this important process. -

Barhl1regulates Migration and Survival of Cerebellar Granule
3104 • The Journal of Neuroscience, March 24, 2004 • 24(12):3104–3114 Development/Plasticity/Repair Barhl1 Regulates Migration and Survival of Cerebellar Granule Cells by Controlling Expression of the Neurotrophin-3 Gene Shengguo Li,1 Feng Qiu,1 Anlong Xu,2 Sandy M. Price,1 and Mengqing Xiang1 1Center for Advanced Biotechnology and Medicine and Department of Pediatrics, University of Medicine and Dentistry of New Jersey-Robert Wood Johnson Medical School, Piscataway, New Jersey 08854, and 2Department of Biochemistry, College of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China The neurons generated at the germinal rhombic lip undergo long distance migration along divergent pathways to settle in widely dispersed locations within the hindbrain, giving rise to cerebellar granule cells and precerebellar nuclei. Neurotrophin-3 (NT-3) signaling has been shown to be required for proper migration and survival of cerebellar granule cells. The molecular bases that govern NT-3 expression within the cerebellum, however, remain unknown at present. Here we report that, during early mouse neurogenesis, the Barhl1 homeobox gene is highly expressed by the rhombic lip and rhombic lip-derived migratory neurons. Its expression is later restricted to cerebellar granule cells and precerebellar neurons extending mossy fibers, two groups of neurons that synaptically connect in the adult cerebellar system. Loss of Barhl1 function causes cerebellar phenotypes with a striking similarity to those of NT-3 conditional null mice, which include attenuated cerebellar foliation as well as defective radial migration and increased apoptotic death of granule cells. Correlating with these defects, we find that NT-3 expression is dramatically downregulated in granule cells of the posterior lobe of Ϫ Ϫ Ϫ Ϫ Barhl1 / cerebella. -

Staining of Cerebellar Cortex Granular Layer Interneurons with Natural Dye of Madder Anneh Mohammad Gharravi
Gharravi Cerebellum & Ataxias (2016) 3:12 DOI 10.1186/s40673-016-0050-6 RESEARCH Open Access Staining of cerebellar cortex granular layer interneurons with natural dye of Madder Anneh Mohammad Gharravi Abstract Background: The objective of the present study was an investigation of root Rubia Tinctorum (Madder) as a natural dye to identification of granular layer interneurons of the rat cerebellum. Methods: Seven to ten micrometre sections were collected from the cerebellum and stained only with Madder for 2, 24 and 48 h. Other sections were stained with Madder then with hematoxyllin, cresyl violet, eosin, light green. Microscopic identification of cells was performed based on cell morphology, reaction and binding of with the dye. All data were expressed as mean ± SD in and significance was set at p ≤0.05. Results: Madder with alum as mordant resulted a deep red staining of interneurons. Unipolar brush cells (UBCs) were observed with a cell body diameter intermediate between granule and Golgi cells in the superficial layer of the granular layer. Golgi cells were identified almost as large as Purkinje cells with irregular rounded or polygonal morphology. Lugaro cells were observed as spindle-shaped cells adjacent to Purkinje layer. Conclusion: Results of the present study showed that mader could stain granular layer interneurons in cerebellum cortex of rat. Keywords: Madder, Cerebellum, Unipolar brush cells, Lugaro cell, Golgi neurons Background with all cerebellar cortical neurons and fibers and they Histologically, the cerebellar cortex is divided into three function as inhibitory interneurons. These spindle-shaped layers: the molecular, the Purkinje and the granular cells locate just underneath the Purkinje cell layer are layers. -

Centrosome Positioning in Vertebrate Development
Commentary 4951 Centrosome positioning in vertebrate development Nan Tang1,2,*,` and Wallace F. Marshall2,` 1Department of Anatomy, Cardiovascular Research Institute, The University of California, San Francisco, USA 2Department Biochemistry and Biophysics, The University of California, San Francisco, USA *Present address: National Institute of Biological Science, Beijing, China `Authors for correspondence ([email protected]; [email protected]) Journal of Cell Science 125, 4951–4961 ß 2012. Published by The Company of Biologists Ltd doi: 10.1242/jcs.038083 Summary The centrosome, a major organizer of microtubules, has important functions in regulating cell shape, polarity, cilia formation and intracellular transport as well as the position of cellular structures, including the mitotic spindle. By means of these activities, centrosomes have important roles during animal development by regulating polarized cell behaviors, such as cell migration or neurite outgrowth, as well as mitotic spindle orientation. In recent years, the pace of discovery regarding the structure and composition of centrosomes has continuously accelerated. At the same time, functional studies have revealed the importance of centrosomes in controlling both morphogenesis and cell fate decision during tissue and organ development. Here, we review examples of centrosome and centriole positioning with a particular emphasis on vertebrate developmental systems, and discuss the roles of centrosome positioning, the cues that determine positioning and the mechanisms by which centrosomes respond to these cues. The studies reviewed here suggest that centrosome functions extend to the development of tissues and organs in vertebrates. Key words: Centrosome, Development, Mitotic spindle orientation Introduction radiating out to the cell cortex (Fig. 2A). In some cases, the The centrosome of animal cells (Fig. -

Control of Cerebellar Granule Cell Output by Sensory-Evoked Golgi Cell Inhibition
Control of cerebellar granule cell output by sensory-evoked Golgi cell inhibition Ian Duguid1,2,3, Tiago Branco1,4, Paul Chadderton5, Charlotte Arlt, Kate Powell6, and Michael Häusser3 Wolfson Institute for Biomedical Research and Department of Neuroscience, Physiology, and Pharmacology, University College London, London WC1E 6BT, United Kingdom Edited by Masao Ito, RIKEN Brain Science Institute, Wako, Japan, and approved September 1, 2015 (received for review May 25, 2015) Classical feed-forward inhibition involves an excitation–inhibition Results sequence that enhances the temporal precision of neuronal re- Sensory-Evoked Phasic and Spillover Golgi Cell Inhibition Precedes sponses by narrowing the window for synaptic integration. In Mossy Fiber Excitation in Granule Cells. Cerebellar granule cells the input layer of the cerebellum, feed-forward inhibition is thought receive direct phasic and indirect or “spillover” GABAergic in- to preserve the temporal fidelity of granule cell spikes during mossy put from Golgi cells (6, 16, 20, 21). To investigate the temporal fiber stimulation. Although this classical feed-forward inhibitory cir- dynamics of sensory-evoked inhibition in vivo, we recorded cuit has been demonstrated in vitro, the extent to which inhibition spontaneous and sensory-evoked excitatory (Vhold = −70 mV) shapes granule cell sensory responses in vivo remains unresolved. and inhibitory (Vhold = 0 mV) currents from the same granule Here we combined whole-cell patch-clamp recordings in vivo and cells in Crus II (Fig. 1 A–D). Granule cells were identified based dynamic clamp recordings in vitro to directly assess the impact of on their characteristic electrophysiological properties (Table S1), Golgi cell inhibition on sensory information transmission in the depth from the pial surface (>250 μm), and morphology (Fig. -

Forward Signaling by Unipolar Brush Cells in the Mouse Cerebellum
Cerebellum DOI 10.1007/s12311-015-0693-5 ORIGINAL PAPER Forward Signaling by Unipolar Brush Cells in the Mouse Cerebellum Stijn van Dorp1 & Chris I. De Zeeuw 1,2 # The Author(s) 2015. This article is published with open access at Springerlink.com Abstract Unipolar brush cells (UBCs) are glutamatergic in- Introduction terneurons prominently present in the granular layer of the vestibulocerebellum. UBCs engage in extensive synaptic con- Forward processing, in particular the absence of recurrent tact with a single presynaptic mossy fiber and signal to down- excitation, is a defining feature of cerebellar architecture stream granule cells through an elaborate network of mossy and computation. In the granular layer of the fiber-like axons. Ultrastructural examinations and electro- vestibulocerebellum, unipolar brush cells (UBCs) provide physiological recordings in organotypic slice cultures have a powerful forward excitatory action onto granule cells indicated that UBCs target not only granule cells but also other through a cortex-intrinsic network of mossy fiber-like UBCs, thus forming chains of two or perhaps more intercon- axons [1, 2]. UBCs are characterized by an elaborate nected UBCs. In this report, we show recordings of spontane- brush-like dendrite (Fig. 1a) that forms an unusually exten- ous and evoked (di)synaptic events in granule cells and UBCs sive synaptic contact with a single presynaptic mossy fiber in fresh cerebellar slices from juvenile mice (5–7 weeks). The rosette [3]. This highly specialized configuration has been patterns of arrival of synaptic events were consistent with the proposed to facilitate prolonged entrapment of glutamate presence of a presynaptic UBC, and recordings from UBCs in the synaptic cleft, underlying complex temporal trans- displayed spontaneous protracted synaptic events characteris- formations of incoming mossy fiber signals [4, 5]. -

Starch Granule Initiation in Arabidopsis Thaliana Chloroplasts
The Plant Journal (2021) doi: 10.1111/tpj.15359 FOCUSED REVIEW Starch granule initiation in Arabidopsis thaliana chloroplasts Angel Merida 1 and Joerg Fettke2,* 1Institute of Plant Biochemistry and Photosynthesis (IBVF), Consejo Superior de Investigaciones Cientıficas (CSIC), Universidad de Sevilla (US), Avda Americo Vespucio, 49, Sevilla 41092, Spain, and 2Biopolymer Analytics, Institute of Biochemistry and Biology, University of Potsdam, Karl-Liebknecht-Str. 24-25, Building 20, Potsdam-Golm 14476, Germany Received 1 April 2021; revised 14 May 2021; accepted 22 May 2021. *For correspondence (e-mail [email protected]). SUMMARY The initiation of starch granule formation and the mechanism controlling the number of granules per plastid have been some of the most elusive aspects of starch metabolism. This review covers the advances made in the study of these processes. The analyses presented herein depict a scenario in which starch synthase isoform 4 (SS4) provides the elongating activity necessary for the initiation of starch granule formation. However, this protein does not act alone; other polypeptides are required for the initiation of an appropriate number of starch granules per chloroplast. The functions of this group of polypeptides include providing suitable substrates (mal- tooligosaccharides) to SS4, the localization of the starch initiation machinery to the thylakoid membranes, and facilitating the correct folding of SS4. The number of starch granules per chloroplast is tightly regulated and depends on the developmental stage of the leaves and their metabolic status. Plastidial phosphorylase (PHS1) and other enzymes play an essential role in this process since they are necessary for the synthesis of the sub- strates used by the initiation machinery. -

The Structure of Plastids and Other Cytoplasmic Bodies in Fixed Preparations of Epidermal Strips
THE STRUCTURE OF PLASTIDS AND OTHER CYTOPLASMIC BODIES IN FIXED PREPARATIONS OF EPIDERMAL STRIPS By J. G. BALD'* (Plate 1) [Manuscript received October 6, 1948] Summary The fixation of the stromatic structure of plastids was found possiblte by the use of mixtures designed for the fixation of viruses in infected plant tissues. Other features of plastids seen in fixed material are described. Bodies formerly assumed to be' protein crystals are fixed in a form that suggests a less simple structure and possibly a more important function than that of reserve protein. At times there seems to be an association between plastids and bodies that is partly depen<}ent on incident light. I. INTRODUCTION The structure of plastids has been discovered mainly from observations on living material (Weier 1938; Jungers and Doutreligne 1943). Most of the estab lished fixatives seriously distort the stroma, and in doing so they destroy the plastids' most characteristic structural feature (Zirkle 1926). During experiments with fixatives intended to facilitate the staining of viruses in infected plant tissues (Bald 1948b ), it was found that the stromatic structure of the plastids was some times preserved. Fixatives were developed that consistently preserve this and possibly other essential features of the plastids. In addition, granules that have been in one of their forms called by virus workers "cuboidal bodies" (Rawlins and Johnson 1925; Goldstein 1926; Holmes 1928; Clinch 1932; Woods 1933) have appeared as portions of composite struc tures that superficially were somewhat like immature plastids. If the whole structures have not previously been observed and described, it is because portions of them are artefacts due to these newly-developed fixatives; or else the more delicate parts are easily destroyed by other types of fixation. -
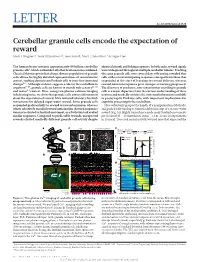
Cerebellar Granule Cells Encode the Expectation of Reward Mark J
LETTER doi:10.1038/nature21726 Cerebellar granule cells encode the expectation of reward Mark J. Wagner1*, Tony Hyun Kim1,2*, Joan Savall1, Mark J. Schnitzer1,3 & Liqun Luo1 The human brain contains approximately 60 billion cerebellar identical stimuli and licking responses. In both tasks, reward signals granule cells1, which outnumber all other brain neurons combined. were widespread throughout multiple cerebellar lobules. Tracking Classical theories posit that a large, diverse population of granule the same granule cells over several days of learning revealed that cells allows for highly detailed representations of sensorimotor cells with reward-anticipating responses emerged from those that context, enabling downstream Purkinje cells to sense fine contextual responded at the start of learning to reward delivery, whereas changes2–6. Although evidence suggests a role for the cerebellum in reward-omission responses grew stronger as learning progressed. cognition7–10, granule cells are known to encode only sensory11–13 The discovery of predictive, non-sensorimotor encoding in granule and motor14 context. Here, using two-photon calcium imaging cells is a major departure from the current understanding of these in behaving mice, we show that granule cells convey information neurons and markedly enriches the contextual information available about the expectation of reward. Mice initiated voluntary forelimb to postsynaptic Purkinje cells, with important implications for movements for delayed sugar-water reward. Some granule cells cognitive processing in the cerebellum. responded preferentially to reward or reward omission, whereas Mice voluntarily grasped the handle of a manipulandum (Methods) others selectively encoded reward anticipation. Reward responses and pushed it forward up to 8 mm for delayed receipt of a sucrose-water were not restricted to forelimb movement, as a Pavlovian task evoked reward (Fig.