Road Kill of Kangaroos on an Australian Outback Highway
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sealing the Cobb and Silver City Highways Community Update April 2020
Transport for NSW Sealing the Cobb and Silver City highways Community update I April 2020 The NSW Government is providing $145 million to rebuild and seal the remaining sections of both the Cobb and Silver City highways, bringing the total invested since 2011 to $195 million. Rebuilding these highways will greatly improve the safety and reliability of routes for trade, tourism and local communities. In December 2020 the Far West Project Team earned the title of Transport for NSW "Project Team of the Year" for their ongoing achievements and commitment to deliver. We asked some of the team: What do you enjoy about working in the Far West? Ethan Degoumois, Anthony Tom Smith, Ben Ragenovich, Tayla Doubtfire, Sabrina Trezise, Road Worker: Campbell, Civil Truck Driver: Safety Civil Construction Road Worker: I enjoy working Construction I was born and Environment and Trainee: Connecting out bush with Trainee: bred in the bush Quality Officer: I like working communities gives a good crew. I like working in and I know the I enjoy the remotely in a me a feeling of Weather can be a new places over importance of isolation the Far construction immense pride. challenge, some the Far West and accessible roads West offers. It environment. I In addition, I would days it can be working with the in the outback. forces us to adapt have also become like to be a role 45°C and the next older generation, I enjoy being and grow the close friends with model for younger it could be raining. learning from the part of the team way we work to the person I live generations, stories they tell. -

Newsletter 112 February 2016
Maltese Newsletter 112 February 2016 We salute the Maltese organizations in South Australia for their sterling work among the members of the Maltese community The Maltese Guild of South Australia The Chaplain Festivities Group The Maltese RSL Sub branch The Maltese Queen of Victories Band The St Catherine Society of SA The Maltese Senior Citizens of SA The Maltese Community Radio EBIfm The Blue Grotto Maltese Program PBAfm The Society of Christian Doctrine The Maltese Aged Care Association of SA Other institutions Consulate for Malta in SA Maltese Chaplaincy Maltese Franciscan Sisters of the Sacred Heart THE MALTESE COMMUNITY COUNCIL www.ozmalta.page4.me/ Page 1 Maltese Newsletter 112 February 2016 MALTESE PEOPLE ARE IN EVERY CORNER OF THE WORLD MALTESE AT BROKEN HILL NSW Broken Hill is an isolated mining city in the far west of outback New South Wales, Australia. The "BH" is the world's largest mining company, BHP Billiton, refers to "Broken Hill" and its early operations in the city. Broken Hill is located near the border with South Australia on the crossing of the Barrier Highway and the Silver City Highway , in the Barrier Range. It is 315 m (1,033 ft) above sea level, with a hot desert climate. The closest major city is Adelaide, the capital of South Australia, which is more than 500 km to the southwest. Broken Hill has been referred to as "The Silver City", the "Oasis of the West", and the "Capital of the Outback” Although over 1,100 km (684 mi) west of Sydney and surrounded by semi-desert, the town has prominent park and garden displays and offers a number of attractions such as the Living Desert Sculptures. -

Government Gazette of 28 September 2012
4043 Government Gazette OF THE STATE OF NEW SOUTH WALES Number 100 Friday, 28 September 2012 Published under authority by the Department of Premier and Cabinet LEGISLATION Online notification of the making of statutory instruments Week beginning 17 September 2012 THE following instruments were officially notified on the NSW legislation website (www.legislation.nsw.gov.au) on the dates indicated: Regulations and other statutory instruments Environmental Planning and Assessment Amendment (Contribution Plans) Regulation 2012 (2012-471) — published LW 21 September 2012 Public Finance and Audit Amendment (Prescribed Audits) Regulation 2012 (2012-472) — published LW 21 September 2012 Road Transport (Safety and Traffic Management) Amendment (Removal of Unattended Vehicles) Regulation 2012 (2012-469) — published LW 21 September 2012 Environmental Planning Instruments Hawkesbury Local Environmental Plan 2012 (2012-470) — published LW 21 September 2012 State Environmental Planning Policy Amendment (Miscellaneous) 2012 (2012-473) — published LW 21 September 2012 4044 OFFICIAL NOTICES 28 September 2012 Assents to Acts ACTS OF PARLIAMENT ASSENTED TO Legislative Assembly Office, Sydney, 24 September 2012 IT is hereby notified, for general information, that Her Excellency the Governor has, in the name and on behalf of Her Majesty, this day assented to the undermentioned Acts passed by the Legislative Assembly and Legislative Council of New South Wales in Parliament assembled, viz.: Act No. 65 2012 – An Act to amend the Classification (Publications, Films and Computer Games) Enforcement Act 1995 to provide for the enforcement of an R 18+ classification category for computer games; and for related purpose. [Classification (Publications, Films and Computer Games) Enforcement Amendment (R18+ Computer Games) Bill] Act No. -

Minutes of the Tourist Attraction Signposting Assessment Committee
TASAC Minutes 20 January 2016 Minutes of the Tourist Attraction Signposting Assessment Committee Wednesday 20 January 2016 at the RMS Parramatta office Level 5, 27-31 Argyle Street Parramatta Members David Douglas Regional Coordinator TASAC and Drive, Destination NSW Phil Oliver Guidance and Delineation Manager, Roads & Maritime Services (RMS) Maria Zannetides TASAC Secretariat Also Present Cameron McIntyre TEO, RMS Sydney Region John Rozos RMS Sydney Region (part meeting) AGENDA ITEMS 1. DELEGATIONS / PRESENTATIONS & REGIONAL SIGNPOSTING ISSUES N / A 2. NEW TOURIST SIGNPOSTING APPLICATIONS 2.1 Paroo Darling National Park, near Wilcannia An application has been lodged to review the eligibility of Paroo Darling National Park for tourist signposting (TASAC found the park to be eligible for signposting in 2008) and allow some of the park’s signage to be updated and also to secure signposting for a new precinct known as Peery Lake Picnic Area within the Paroo Darling Overflow Section of the park. The park is in the north west corner of the State, north east of the Cobb Highway and north of the Barrier Highway. The nearest towns are Wilcannia and White Cliffs, both to the west of the park. The park conserves extensive semi-permanent freshwater wetlands associated with both the Paroo and Darling Rivers. The area is internationally significant for bird migration and recognised under the Ramsar Treaty for conserving wetlands of international importance. Additionally, Peery Lake is the only lake bed in the Southern Hemisphere where mound springs (natural outlets for artesian water) are located. Various Aboriginal artefacts and sites exist in the area and the lake has been recorded as a tourist attraction since the 1910s. -

Sealing of the Silver City Highway Broken Hill to Tibooburra Commemorative Community Update Connecting the Corner Country Reopened on Wednesday 1 July, 2020
Sealing of the Silver City Highway Broken Hill to Tibooburra Commemorative Community Update Connecting the Corner Country Reopened on Wednesday 1 July, 2020 Transport for NSW project team practicing physical distancing on the Silver City Highway The NSW Government committed $145 million to rebuild and seal the remaining sections of the Cobb and Silver City highways. Today we commemorate the completion of the works on the Silver City Highway between Broken Hill and Tibooburra, proudly delivered earlier than scheduled, improving freight access, flood immunity, regional development and road safety. The history The Silver City Highway connects Buronga, on the Victorian border with the Queensland border via Broken Hill and Tibooburra. The first motorised vehicles started travelling between Broken Hill and Tibooburra in the 1920s. It is said to have taken three days as they followed the tracks that were formed by horses and camels, repeatedly getting bogged in sand hills. Travelling via camel took eight days, which meant the reduction to three was a significant improvement at the time. The upgrade works have seen the existing road rebuilt to include table drains on both sides of the road, eliminating pooling rain water, resulting in a dramatic decrease in closures each year, more reliable access and a smoother journey. Now 100 years on we can travel between these remote locations in just over 3.5 hours. The journey 10k WARRI GATE 53.1 kilometres of unsealed road remaining Tibooburra to Warri Gate Surface preparation, Eight Mile, Silver City -

Community Update Murray to Broken Hill Pipeline June 2017
Community Update Wentworth Murray to Broken Hill pipeline June 2017 Project background How will the pipeline impact the WaterNSW is delivering the Lower Darling River? 270km Murray to Broken Hill The NSW Government is delivering a range of projects to pipeline – the NSW improve the management of the Darling River. Those Government’s largest projects, and decisions about future operation of the Darling investment in regional water River, are managed by the NSW Government. security. This project will provide the much needed long term solution to secure the water supply for Broken Hill. To Broken Hill Silver City Highway Proposed route near Pipeline Sheoak Lane Wentworth Consultation with stakeholders, cultural and Renmark Road Pooncarie Road environmental assessments and technical requirements have informed the proposed ek route of the pipeline around the eastern Boydes Cre side of Wentworth. The proposed route is in road reserves where possible and runs along the eastern side of the levee bank. Wentworth Street Smyth Street The facts about Darling Street the pipeline d a o R Pipeline Pavilion Road th D r • The water will be drawn from the River o a w r t Wentworth l n i n e Silver City Highway (Adams Street) Murray adjacent to the existing Council g W d l R O i v water extraction facility on the eastern e r Emily Street side of Wentworth William Street • The pipeline will supply up to 37.4 ek megalitres of raw water to Essential e r C s r Water in Broken Hill Armstrong Avenue e k c u T • The route will follow the Silver City Highway corridor to Broken Hill • The pipeline will be underground Delta Road d a H o • It will operate most of the time, except ospital R Silver City Highway during routine maintenance activities Pipeline • Construction of the pipeline is expected to generate up to 200 jobs Ski Reserve Road Offtake y urra er M Riv What about the Menindee Lakes? Broken Hill’s drinking water requirement (≈10GL), is and SA, as signatories of the Murray-Darling small in comparison to other water allocations (South Basin Agreement. -

Stay in the Outback with Friends
Mutawintji National Park Paroo-Darling National Park HIGHWAY Cobar Florida BARRIER Wilcannia Wongalara A32 ROUTEHermidale MAP #4 HIGHWAY Poopelloe Lake Canbelego Lake Emmdale 2019 edition A32 A32 Roadhouse To Sydney Stephens Creek Kidman Little Topar Creek BARRIER NELIA River COBB Silverton Roadhouse GAARI Broken Hill 8 S A32 I L V Nymagee Mingary E Cockburn 117 Buckwarooh R Darling Pamamaroo B87 To Adelaide C Lake Outback I Side trip to B75 T Sunset Strip Way Y Broken Hill 8 Beds 85 Menindee FOOD ANDWINETOURINGROUTE B79 H Lake 117 Menindee IG Ratcatchers H Kinchega Lake HIGHWAY W Victoria A National Park Yathong Y Lake Lake 55 Kajuligah Nature Reserve Tandou Nature Res Gum Lake 47 28 IVANHOE Tallebung Sayers Lake CARAVAN BINDARA on OUTBACK BEDS Darnick Mount Hope the DARLING PARK Beilpajah Ivanhoe Round Hill Conoble NR Euabalong Coombah WALES Moornanyah Trida Roadhouse Lake Roto West B75 Way Euabalong NEW SOUTH Willandra Popiltah 97 Mulurulu National Park Danggali Lake Murrin Bridge Cons Park Lake Mossgiel intheOutback “...stay 8 COBB Lake Cargelligo Travellers Wallanthery Pooncarie Lake Garnpung with Friends...” Tullibigeal Lake Mungo HIGHWAY Kidman Lake Tarawi 27 Brewster Side trip to National Park Hillston Nature Res Wentworth Top B87 & Mildura River Hut Rd To Sydney Kikoira Kidman Lake Rankins Darling Mungo River Springs Yalgogrin Merriwagga HWY Weethalle 120 Hatfield Booligal 91 VICTORIA B64 Lake WESTERN Victoria Goolgowi Way Buralyang Tallimba 8 10 Gunbar Cocoparra Dareton 3 B75 Wentworth 2 Mallee Cliffs Tabbita National Park Lindsay -

Filling the Void: Silver City Highway
Proceedings of the 21st Association of Public Authority Surveyors Conference (APAS2016) Leura, New South Wales, Australia, 4-6 April 2016 Filling the Void: Silver City Highway Greg Goodman LandTeam [email protected] ABSTRACT ‘Filling the Void’ was a collaborative LandTeam Australia Pty Ltd / Roads and Maritime Services (RMS) submission into the 2015 Excellence in Surveying and Spatial Information Awards. Having been born off the land in the southwest of New South Wales and having been fortunate enough in the 1970s to travel as a surveyor to remote areas of Australia with BHP, to say that the author has developed an affinity with the outback is an understatement – I simply love it! This passion for the arid zone has been fuelled over the years with regular non-surveying trips to inland Australia and its bewildering environs. So, when an opportunity arose in June 2014 to perform the control and detail surveys for an RMS upgrade of a remote 14 km section of the Silver City Highway some 230 km north of Broken Hill, the excitement was difficult to contain – the opportunity to spend perhaps 2 weeks getting to know intimately a ‘micro section’ of the outback whilst working is a rarity. The purpose of the survey brief was to procure a detail engineering survey with network control suitable for input into detail road design and network control for the HW22 Shannons Creek Initial Sealing Project and designed to allow the Royal Flying Doctor Service to land on the highway to deal with emergency medical matters within the wide environs of the locality. -
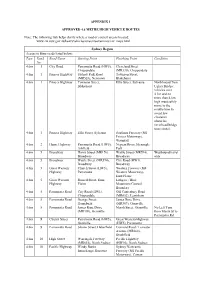
APPENDIX 1 APPROVED 4.6 METRE HIGH VEHICLE ROUTES Note: The
APPENDIX 1 APPROVED 4.6 METRE HIGH VEHICLE ROUTES Note: The following link helps clarify where a road or council area is located: www.rta.nsw.gov.au/heavyvehicles/oversizeovermass/rav_maps.html Sydney Region Access to State roads listed below: Type Road Road Name Starting Point Finishing Point Condition No 4.6m 1 City Road Parramatta Road (HW5), Cleveland Street Chippendale (MR330), Chippendale 4.6m 1 Princes Highway Sydney Park Road Townson Street, (MR528), Newtown Blakehurst 4.6m 1 Princes Highway Townson Street, Ellis Street, Sylvania Northbound Tom Blakehurst Ugly's Bridge: vehicles over 4.3m and no more than 4.6m high must safely move to the middle lane to avoid low clearance obstacles (overhead bridge truss struts). 4.6m 1 Princes Highway Ellis Street, Sylvania Southern Freeway (M1 Princes Motorway), Waterfall 4.6m 2 Hume Highway Parramatta Road (HW5), Nepean River, Menangle Ashfield Park 4.6m 5 Broadway Harris Street (MR170), Wattle Street (MR594), Westbound travel Broadway Broadway only 4.6m 5 Broadway Wattle Street (MR594), City Road (HW1), Broadway Broadway 4.6m 5 Great Western Church Street (HW5), Western Freeway (M4 Highway Parramatta Western Motorway), Emu Plains 4.6m 5 Great Western Russell Street, Emu Lithgow / Blue Highway Plains Mountains Council Boundary 4.6m 5 Parramatta Road City Road (HW1), Old Canterbury Road Chippendale (MR652), Lewisham 4.6m 5 Parramatta Road George Street, James Ruse Drive Homebush (MR309), Granville 4.6m 5 Parramatta Road James Ruse Drive Marsh Street, Granville No Left Turn (MR309), Granville -

2014 Conference Minutes
DRAFT WESTERN DIVISION COUNCILS OF NSW March 2- 4, 2014 Hosted by Carrathool Shire Council Western Division Councils Conference 1 EXECUTIVE 2012/2013 President – Councillor Peter Laird Mayor Carrathool Shire Council Vice President- Councillor Ray Longfellow Mayor Central Darling Shire Council Executive - Councillor Darriea Turley Deputy Mayor Broken Hill City Council Councillor Bill Murray Mayor Walgett Shire Council Ruth Fagan -Executive Officer Apologies Parliamentarians: Kevin Humphries, Minister for Western NSW, Member for Barwon, Prue Goward, NSW Minister for Community Services, John Williams, MP Member for Murray Darling, Minister for Primary Industries, Katrina Hodgkinson, Adrian Piccoli, MP Minister for Education, Sussan Ley, MP Federal Member for Farrer, Assistant Minister for Education, Senator Fiona Nash, Assistant Minister for Health, Michael McCormack MP, Member for Riverina OTHERS Steve Toms, Cross Border Commissioner, Craig Knowles, Chair Murray Darling Basin Authority, Geoff McKechnie, Assistant Commission NSW Police Force, Scott McLachlan Chief Executive Western Local Health District, Michael Kneipp, Director, Catchment and Lands, NSW Trade and Investment, Mr Ivor Frischknecht, Australian Renewable Energy Agency, Ross O’Shea District Director Far West Department of Family and Community Services, Mark Peacock – Director Western Branch NPWS, Cr Bill Murray, Mayor Walgett Shire Council, Cr Don McKinnon, Mayor Wentworth Council, Cr Steve O’Halloran, Mayor Balranald Council, Cr Ray Donald, Mayor Bogan Council, Cr Marsha Isbister, -

Chapter 14 Traffic and Transport
Environmental Assessment Report Chapter 14 Traffic and Transport CHAPTER 14 TRAFFIC AND TRANSPORT TABLE OF CONTENTS 14 TRAFFIC AND TRANSPORT............................................................................. 14-1 14.1 Introduction.............................................................................................. 14-1 14.2 Methodology ............................................................................................ 14-1 14.3 Existing Environment.............................................................................. 14-1 14.3.1 Internal roads and car parking...................................................... 14-1 14.3.2 Site access .................................................................................. 14-2 14.3.3 Regional road network ................................................................. 14-2 14.3.4 Local road network....................................................................... 14-3 14.3.5 Rail network ................................................................................. 14-4 14.4 Impact Assessment................................................................................. 14-6 14.4.1 Internal roadways and parking ..................................................... 14-6 14.4.2 Site access .................................................................................. 14-7 14.4.3 Traffic generation on external roads............................................. 14-7 14.4.4 Transport of crushed ore........................................................... -

New South Wales Victoria
PARA OAD NCARIE R DARLING RIVER POO POONCARIE ARUMPO WENTWORTH ARUMPO ROAD Proposal study area Proposal locality (10km buffer) Existing transmission line EUSTON infrastructure COOMEALLA )" BURONGA Buronga substation DARETON 220KV ") SILVER C Red Cliffs substation ITY Major road HIG ") HW Minor road CURLWAA AY Major river MOURQUONG ")") XW Waterbody Bionet threatened fauna records within BOEILL BURONGA Threatened Freshwater Fish 10km CREEK GOL ^_ Habitat (Source: DPIE, 2020) GOL Important Wetlands !( ") ")!( ") Australasian Bittern ^_ ' NPWS reserve GF XW Australian Painted Snipe Black-eared Miner %2%2 MALLEE CLIFFS ^_ WSP field verified threatened %2 MALLEE flora %2 NATIONAL PARK Corben's Long-eared Bat ") ^_ %2 ^_ Koala ") Santalum murrayanum ") ' Malleefowl ^_ Bionet threatened flora Red-tailed Phascogale %2 GF records within 10km %2 Regent Parrot (eastern %2 %,") ") subspecies) !( TRENTHAM XW Southern Bell Frog ") Austrostipa metatoris RED CLIFFS CLIFFS *# Spotted-tailed Quoll %, Solanum karsense 220KV MONAK PARINGI Swift Parrot Swainsona pyrophila VICTORIA ") ST URT NEW SOUTH H ^_ IG HW AY WALES MURRAY RIVER Map: PS117658_EPBC_002_A2_EPBClist Date: 18/08/2020 GF Source: Esri, Maxar, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community Sources: Esri, HERE, Garmin, Intermap, increment P Corp., GEBCO, USGS, FAO, NPS, NRCAN, GeoBase, GF © WSP Australia Pty Limited (WSP) Copyright in the drawings, information and data recorded is the property of WSP. This document and the information are solely for the use of the authorised recipient and this document may not be used, copied or reproduced in whole or part for any purpose other than that which it was supplied by WSP.