Spacing System of the Mysore Slender Loris (Loris Lydekkerianus Lydekkerianus)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rediscovery of Nycticebus Coucang Insularis Robinson, 1917
Sains Malaysiana 47(10)(2018): 2533–2542 http://dx.doi.org/10.17576/jsm-2018-4710-30 Rediscovery of Nycticebus coucang insularis Robinson, 1917 (Primates: Lorisidae) at Tioman Island and its Mitochondrial Genetic Assessment (Penemuan Semula Nycticebus coucang insularis Robinson, 1917 (Primate: Lorisidae) di Pulau Tioman dan Penilaian Genetik Mitokondrianya) JEFFRINE J. ROVIE-RYAN*, MILLAWATI GANI, HAN MING GAN, GILMOORE G. BOLONGON, TAN CHENG CHENG, NORAZLINDA RAZAK, NORSYAMIMI ROSLI, MOHD AZIZOL AZIZ & KALIP MATKASIM ABSTRACT Slow lorises (Nycticebus) consist of eight species native to Southeast Asia while three species are recognised in Malaysia - N. coucang, N. menagensis and N. kayan. This study reports on the rediscovery of the subspecies N. coucang insularis Robinson, 1917 in Tioman Island and the genetic assessment of its mitochondrial DNA variation. Morphological measurements conform the specimen as the putative N. coucang but with distinct colour and markings. Two mitochondrial DNA segments (cytochrome b and control region) were produced from the subspecies representing their first registered sequences in GenBank. Genetically, the subspecies showed 99% of nucleotide similarity to N. coucang species type for both the DNA segments and constitute its own unique haplotype. Phylogenetic trees constructed using three methods (neighbour joining, maximum likelihood and Bayesian inference) showed two major groups within Nycticebus; the basal group was formed by N. pygmaeus while the second group consisted of the remaining Nycticebus species. The phylogenetic position of the subspecies, however, remains unresolved due to the observed mixing between N. coucang and N. bengalensis. Several reasons could lead to this condition including the lack of well documented voucher specimens and the short DNA fragments used. -

Radar.Brookes.Ac.Uk/Radar/Items/C664d3c7-1663-4D34-Aed5-E3e070598081/1
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Oxford Brookes University: RADAR RADAR Research Archive and Digital Asset Repository Pozzi, L, Nekaris, K, Perkin, A, Bearder, S, Pimley, E, Schulze, H, Streicher, U, Nadler, T, Kitchener, A, Zischler, H, Zinner, D and Roos, C Remarkable Ancient Divergences Amongst Neglected Lorisiform Primates Pozzi, L, Nekaris, K, Perkin, A, Bearder, S, Pimley, E, Schulze, H, Streicher, U, Nadler, T, Kitchener, A, Zischler, H, Zinner, D and Roos, C (2015) Remarkable Ancient Divergences Amongst Neglected Lorisiform Primates. Zoological Journal of the Linnean Society, 175 (3). pp. 661-674. doi: 10.1111/zoj.12286 This version is available: https://radar.brookes.ac.uk/radar/items/c664d3c7-1663-4d34-aed5-e3e070598081/1/ Available on RADAR: August 2016 Copyright © and Moral Rights are retained by the author(s) and/ or other copyright owners. A copy can be downloaded for personal non-commercial research or study, without prior permission or charge. This item cannot be reproduced or quoted extensively from without first obtaining permission in writing from the copyright holder(s). The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the copyright holders. This document is the published version of the journal article. WWW.BROOKES.AC.UK/GO/RADAR bs_bs_banner Zoological Journal of the Linnean Society, 2015, 175, 661–674. With 3 figures Remarkable ancient divergences amongst neglected lorisiform primates LUCA -

In Situ Conservation
NEWSN°17/DECEMBER 2020 Editorial IN SITU CONSERVATION One effect from 2020 is for sure: Uncertainty. Forward planning is largely News from the Little Fireface First, our annual SLOW event was impossible. We are acting and reacting Project, Java, Indonesia celebrated world-wide, including along the current situation caused by the By Prof K.A.I. Nekaris, MA, PhD by project partners Kukang Rescue Covid-19 pandemic. All zoos are struggling Director of the Little Fireface Project Program Sumatra, EAST Vietnam, Love economically after (and still ongoing) Wildlife Thailand, NE India Primate temporary closures and restricted business. The Little Fireface Project team has Investments in development are postponed Centre India, and the Bangladesh Slow at least. Each budget must be reviewed. been busy! Despite COVID we have Loris Project, to name a few. The end In the last newsletter we mentioned not been able to keep up with our wild of the week resulted in a loris virtual to forget about the support of the in situ radio collared slow lorises, including conference, featuring speakers from conservation efforts. Some of these under welcoming many new babies into the the helm of the Prosimian TAG are crucial 11 loris range countries. Over 200 for the survival of species – and for a more family. The ‘cover photo’ you see here people registered, and via Facebook sustainable life for the people involved in is Smol – the daughter of Lupak – and Live, more than 6000 people watched rd some of the poorest countries in the world. is our first 3 generation birth! Having the event. -

World's Most Endangered Primates
Primates in Peril The World’s 25 Most Endangered Primates 2016–2018 Edited by Christoph Schwitzer, Russell A. Mittermeier, Anthony B. Rylands, Federica Chiozza, Elizabeth A. Williamson, Elizabeth J. Macfie, Janette Wallis and Alison Cotton Illustrations by Stephen D. Nash IUCN SSC Primate Specialist Group (PSG) International Primatological Society (IPS) Conservation International (CI) Bristol Zoological Society (BZS) Published by: IUCN SSC Primate Specialist Group (PSG), International Primatological Society (IPS), Conservation International (CI), Bristol Zoological Society (BZS) Copyright: ©2017 Conservation International All rights reserved. No part of this report may be reproduced in any form or by any means without permission in writing from the publisher. Inquiries to the publisher should be directed to the following address: Russell A. Mittermeier, Chair, IUCN SSC Primate Specialist Group, Conservation International, 2011 Crystal Drive, Suite 500, Arlington, VA 22202, USA. Citation (report): Schwitzer, C., Mittermeier, R.A., Rylands, A.B., Chiozza, F., Williamson, E.A., Macfie, E.J., Wallis, J. and Cotton, A. (eds.). 2017. Primates in Peril: The World’s 25 Most Endangered Primates 2016–2018. IUCN SSC Primate Specialist Group (PSG), International Primatological Society (IPS), Conservation International (CI), and Bristol Zoological Society, Arlington, VA. 99 pp. Citation (species): Salmona, J., Patel, E.R., Chikhi, L. and Banks, M.A. 2017. Propithecus perrieri (Lavauden, 1931). In: C. Schwitzer, R.A. Mittermeier, A.B. Rylands, F. Chiozza, E.A. Williamson, E.J. Macfie, J. Wallis and A. Cotton (eds.), Primates in Peril: The World’s 25 Most Endangered Primates 2016–2018, pp. 40-43. IUCN SSC Primate Specialist Group (PSG), International Primatological Society (IPS), Conservation International (CI), and Bristol Zoological Society, Arlington, VA. -

SILVERY GIBBON PROJECT Newsletterthe Page 1 September 2013 SILVERY GIBBON PROJECT
SILVERY GIBBON PROJECT NEWSLETTERThe Page 1 September 2013 SILVERY GIBBON PROJECT PO BOX 335 COMO 6952 WESTERN AUSTRALIA Website: www.silvery.org.au E-mail: [email protected] Phone: 0438992325 September 2013 Agile Gibbons, Siamangs, Sunbears, Clouded Leopards and even Sumatran Tigers, the area lies PRESIDENT’S REPORT adjacent to a larger protected forest and is also the location of Kalaweit Conservation Centre. We Dear Members and Friends hope to be able to provide additional support to this important project into the future. Keep an eye We are excited to report that the release of on our Facebook page for updates on camera trap Sadewa and Kiki in June went very well and they images from Supayang. continue to thrive in the forest. They are proving a little challenging for the monitoring team to keep Back in Perth, the SGP team is gearing up for our up with but thanks to their morning call the team Art Auction which will be held on October 26. are still able to locate them most days. Please Once again we have secured some amazing read the update on their release on page 4. pieces and the generosity of artists is truly inspiring. Be sure to get your tickets early as this event may sell out. We also have some exciting projects underway with Wildlife Asia. You can find out more about our crowd funding project on page 5. It was certainly an honour to be present at the release, which was the culmination of many years hard work for the Java Gibbon Centre (JGC) team. -

Infant Development in the Slender Loris (Loris Lydekkerianus Lydekkerianus)
RESEARCH ARTICLES Infant development in the slender loris (Loris lydekkerianus lydekkerianus) Sindhu Radhakrishna1,* and Mewa Singh Department of Psychology, University of Mysore, Mysore 570 006, India 1Present address: National Institute of Advanced Studies, Indian Institute of Science Campus, Bangalore 560 012, India lorises and bushbabies)1, the lorids (angwantibos, pottos In this article we present data on infant development in wild slender loris, a nocturnal primate species. The and lorises) typically have lower developmental and repro- ductive rates in comparison to similar-sized galagos (bush- behavioural ecology of the grey slender loris Loris lyde- 2–4 kkerianus lydekkerianus, a nocturnal strepsirrhine, babies) . Litter size in lorids and galagos appears to be was studied for 21 months (October 1997–June 1999) linked to the stability of the habitat more than body size: in a scrub jungle in Dindigul, south India. A total of species occupying harsher environments tend to have 22,834 scans were collected during 2656 h of observa- higher reproductive rates3,5. Strepsirrhine neonates are tion on identified and unidentified lorises using in- carried for short periods following birth and mode of stantaneous point and ad libitum sampling methods. infant transport can be oral (Galagos bushbabies, Cheiro- Developmental schedules were observed for twelve in- galeus dwarf lemurs, Microcebus mouse lemurs) or the dividuals born during the course of the study period. abdominal fur of the mother (Loris slender loris)3,6,7. In- A greater number of twin births were observed than fant parking is typical of the Loriformes, wherein infants singleton births and more isosexuals than heterosexu- als. -

Hair Morphology of Gray Slender Loris (Loris Lydekkerianus) and Bengal
Journal of Entomology and Zoology Studies 2017; 5(3): 386-388 E-ISSN: 2320-7078 P-ISSN: 2349-6800 Hair morphology of Gray Slender Loris (Loris JEZS 2017; 5(3): 386-388 © 2017 JEZS lydekkerianus) and Bengal Slow Loris (Nycticebus Received: 28-03-2017 Accepted: 29-04-2017 bengalensis) Manokaran Kamalakannan Zoological Survey of India, M-Block, New Alipore, Kolkata, Manokaran Kamalakannan West Bengal, India Abstract The present study was undertaken to know the hair morphology of two Indian lorises Gray Slender Loris Loris lydekkerianus and Bengal Slow Loris Nycticebus bengalensis. The laboratory study was conducted at the Mammal & Osteology section, Zoological Survey of India, Kolkata during January – July 2016. Microscopic hair analysis of two lorises was done using an optical light microscope for its species identification. The hair of two lorises can easily be identified on basis of their ‘unicellular regular’ and ‘ladder’ structure of medulla and ‘oval or circular’ shape of the cross-section of hair. The high-resolution photo-micrographs and key characteristics of hair presented here can be used as an appropriate reference for species identification. Keywords: Lorises, hair morphology, dorsal guard hair, unicellular regular, ladder 1. Introduction The identification of mammal species through hair morphology is one of the methods adapted for identification of confiscated materials received from the different wildlife enforcement agencies and the analysis of mammal hairs has long been useful in the study of prey-predator food habits using the scat-hair analysis [3, 9, 10]. Adequate and systematic knowledge of the structure of dorsal guard hair is necessary to identify the species and data generated from microscopic characteristics of dorsal guard hair will provide for a preparation of identification keys for respective species [4]. -
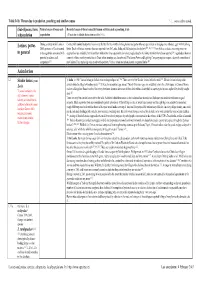
Table 14 B: Threat Due to Predation, Poaching and Similar Causes 1, 2,
Table 14 b: Threat due to predation, poaching and similar causes 1, 2, ... : source, author quoted. (Sub-)Species, form, Natural causes of losses such Recorded causes of threat caused by human activities such as poaching, trade subpopulation as predation (Threat due to habitat destruction see table 14 c) Lorises, pottos During a survey in Sri Lanka in Contact with uninsulated power lines is may be fatal for the slowly climbing lorises and pottos who are specialized on bridging over substrate gaps with their long 2001, presence of Loris seemed limbs. Death of lorises on power lines are reported from Sri Lanka, India and Malaysia (see also below) 207, 66, 211. From Africa, no data concerning pottos or in general to be negatively associated with angwantibos are available, but the problem with power lines apparently also exists, regular deaths of colobus monkeys have been reported 208, regarded as the main potential predators and cause of colobus monkey mortalities in Diani; other monkeys are also affected. The Kenya Power and Lighting Company engineers spent a day with coworkers of competitors 211. the Colobus Trust exploring ways to solve the problem. Victims of electrocution showed amputated limbs 209. Asian lorises L I Slender lorises, genus In India, in 1981 "lorises belong to India's most endangered species" 116. There are very few Slender lorises left in the wild 121. Slender lorises belong to the Loris animals found in illegal ownership (pets) 110. It was also a popular cage animal. Twenty-five years ago, one could buy a loris for a few rupees in Chennai Moore market or Bangalore Russel market. -

Groves #3 Layout 1
Vietnamese Journal of Primatology (2009) 3, 37-44 Diet and feeding behaviour of pygmy lorises (Nycticebus pygmaeus) in Vietnam Ulrike Streicher Wildlife Veterinarinan, Danang, Vietnam. <[email protected]> Key words: Diet, feeding behaviour, pygmy loris Summary Little is known about the diet and feeding behaviour of the pygmy loris. Within the Lorisidae there are faunivorous and frugivorous species represented and this study aimed to characterize where the pygmy loris (Nycticebus pygmaeus) ranges on this scale. Feeding behaviour was observed in adult animals which had been confiscated from the illegal wildlife trade and housed at the Endangered Primate Rescue Center at Cuc Phuong National Park for some time before they were radio collared and released into Cuc Phuong National Park. The lorises were located in daytime by methods of radio tracking and in the evenings they were directly observed with the help of red-light torches. The observed lorises exploited a large variety of different food sources, consuming insects as well as gum and other plant exudates, thus appearing to be truly omnivorous. Seasonal variations in food preferences were observed. Omnivory can be an adaptive strategy, helping to overcome difficulties in times of food shortage. The pygmy loris’ feeding behaviour enables it to rely on other food sources like gum in times when other feeding resource become rare. Gum as an alternative food sources has the advantage of being readily available all year round. However it does not permit the same energetic benefits and consequently the same lifestyle as other food sources. But it is an important part of the pygmy loris’ multifaceted strategy to survive times of hostile environmental conditions. -

Loris Husbandry Manual REPRODUCTION Determining
Loris Husbandry Manual REPRODUCTION Determining Gender Slender Loris The penis and clitoris of lorises are similar in size, shape, and color, which complicates gender determination. The slender lorises can be even harder to sex because their testes are sometimes partially or completely inguinal. The size and location of their testes vary. The vagina is often covered with fur in young females, and close examination is often necessary for safe determination. The tip of the slender loris’ clitoris shows a gray patch (Meier et al., in preparation). The clitoris is smooth with the exception of hairs on the tip, as shown in Figure 13. The penis is covered with fine, velvet-like hairs that are barely visible. Figure 13: Discrimination of sex in lorises 28 Reproduction Slow and Pygmy Loris The penis of the slow and pygmy loris is shorter and thicker than the slender’s penis. The baculum is more developed and the callous pad at the tip of the baculum is larger. The scrotum differs from the slender loris’ scrotum by a distinct median ridge. The clitoris of the slow and pygmy loris is similar to the slender loris’ clitoris but it is relatively shorter and much thicker. The urethra opens at the clitoris tip. The vaginal opening is V-shaped and is situated at the root of the clitoris. Sex determination can often be accomplished shortly after birth in slow and pygmy lorises. The clitoris has a vertical slit at the tip, whereas the penis tip is more blunt and the slit is absent. In adults, the male genitalia show seasonal changes, and testes increase in size during late summer. -

Javan Slow Loris Nycticebus Javanicus) in a Montane Rainforest
Vol. 24: 95–103, 2014 ENDANGERED SPECIES RESEARCH Published online May 8 doi: 10.3354/esr00585 Endang Species Res Contribution to the Theme Section ‘Conservation and ecology of slow lorises’ FREEREE ACCESSCCESS Densities, distribution and detectability of a small nocturnal primate (Javan slow loris Nycticebus javanicus) in a montane rainforest K. Anne-Isola Nekaris1, Jarot Arisona Aji Pambudi2, Dwi Susanto2, Ridwan Djaffar Ahmad1,2, Vincent Nijman1,* 1Noctural Primate Research Group, Oxford Brookes University, OX3 0BP Oxford, UK 2Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Indonesia, Depok 16424, Indonesia ABSTRACT: Nocturnal mammals can be challenging to survey and, especially for many species that live in dense forest habitats, limited information is available on densities and distributions. We surveyed the endemic Javan slow loris Nycticebus javanicus in the montane forests of Mount Gede Pangrango, West Java, Indonesia. Surveys were conducted on 23 transects (260 h covering some 93 km) walking at variable speeds between 200 and 800 m h−1. Densities on individual tran- sects varied from 0 to 52 ind. km−2, with an overall density of 15.6 ind. km−2 (95% CI 9.7 to 25.2 ind. km−2). Encounter rates per kilometre were strongly influenced by the speed at which transects were walked, with fewer lorises detected at higher speeds. This effect was absent when consider- ing encounter rates per hour. Detectability and behavior of Javan slow lorises were not affected by the amount of lunar light and, in contrast to studies of some of their congeners, we found no evi- dence of lunar phobia or lunar philia. -

An Assessment of Trade in Gibbons and Orang-Utans in Sumatra, Indoesia
AN ASSESSMENT OF TRADE IN GIBBONS AND ORANG-UTANS IN SUMATRA, INDONESIA VINCENT NIJMAN A TRAFFIC SOUTHEAST ASIA REPORT Published by TRAFFIC Southeast Asia, Petaling Jaya, Selangor, Malaysia © 2009 TRAFFIC Southeast Asia All rights reserved. All material appearing in this publication is copyrighted and may be reproduced with permission. Any reproduction in full or in part of this publication must credit TRAFFIC Southeast Asia as the copyright owner. The views of the authors expressed in this publication do not necessarily reflect those of the TRAFFIC Network, WWF or IUCN. The designations of geographical entities in this publication, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of TRAFFIC or its supporting organizations concerning the legal status of any country, territory, or area, or its authorities, or concerning the delimitation of its frontiers or boundaries. The TRAFFIC symbol copyright and Registered Trademark ownership is held by WWF. TRAFFIC is a joint programme of WWF and IUCN. Layout by Noorainie Awang Anak, TRAFFIC Southeast Asia Suggested citation: Vincent Nijman (2009). An assessment of trade in gibbons and orang-utans in Sumatra, Indonesia TRAFFIC Southeast Asia, Petaling Jaya, Selangor, Malaysia ISBN 9789833393244 Cover: A Sumatran Orang-utan, confiscated in Aceh, stares through the bars of its cage Photograph credit: Chris R. Shepherd/TRAFFIC Southeast Asia An assessment of trade in gibbons and orang-utans in Sumatra, Indonesia Vincent Nijman Cho-fui Yang Martinez