Antipredator Behavior and Breeding Success in Greater Golden-Plover and Eurasian Dotterel’
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Migratory Birds Index
CAFF Assessment Series Report September 2015 Arctic Species Trend Index: Migratory Birds Index ARCTIC COUNCIL Acknowledgements CAFF Designated Agencies: • Norwegian Environment Agency, Trondheim, Norway • Environment Canada, Ottawa, Canada • Faroese Museum of Natural History, Tórshavn, Faroe Islands (Kingdom of Denmark) • Finnish Ministry of the Environment, Helsinki, Finland • Icelandic Institute of Natural History, Reykjavik, Iceland • Ministry of Foreign Affairs, Greenland • Russian Federation Ministry of Natural Resources, Moscow, Russia • Swedish Environmental Protection Agency, Stockholm, Sweden • United States Department of the Interior, Fish and Wildlife Service, Anchorage, Alaska CAFF Permanent Participant Organizations: • Aleut International Association (AIA) • Arctic Athabaskan Council (AAC) • Gwich’in Council International (GCI) • Inuit Circumpolar Council (ICC) • Russian Indigenous Peoples of the North (RAIPON) • Saami Council This publication should be cited as: Deinet, S., Zöckler, C., Jacoby, D., Tresize, E., Marconi, V., McRae, L., Svobods, M., & Barry, T. (2015). The Arctic Species Trend Index: Migratory Birds Index. Conservation of Arctic Flora and Fauna, Akureyri, Iceland. ISBN: 978-9935-431-44-8 Cover photo: Arctic tern. Photo: Mark Medcalf/Shutterstock.com Back cover: Red knot. Photo: USFWS/Flickr Design and layout: Courtney Price For more information please contact: CAFF International Secretariat Borgir, Nordurslod 600 Akureyri, Iceland Phone: +354 462-3350 Fax: +354 462-3390 Email: [email protected] Internet: www.caff.is This report was commissioned and funded by the Conservation of Arctic Flora and Fauna (CAFF), the Biodiversity Working Group of the Arctic Council. Additional funding was provided by WWF International, the Zoological Society of London (ZSL) and the Convention on Migratory Species (CMS). The views expressed in this report are the responsibility of the authors and do not necessarily reflect the views of the Arctic Council or its members. -

EUROPEAN BIRDS of CONSERVATION CONCERN Populations, Trends and National Responsibilities
EUROPEAN BIRDS OF CONSERVATION CONCERN Populations, trends and national responsibilities COMPILED BY ANNA STANEVA AND IAN BURFIELD WITH SPONSORSHIP FROM CONTENTS Introduction 4 86 ITALY References 9 89 KOSOVO ALBANIA 10 92 LATVIA ANDORRA 14 95 LIECHTENSTEIN ARMENIA 16 97 LITHUANIA AUSTRIA 19 100 LUXEMBOURG AZERBAIJAN 22 102 MACEDONIA BELARUS 26 105 MALTA BELGIUM 29 107 MOLDOVA BOSNIA AND HERZEGOVINA 32 110 MONTENEGRO BULGARIA 35 113 NETHERLANDS CROATIA 39 116 NORWAY CYPRUS 42 119 POLAND CZECH REPUBLIC 45 122 PORTUGAL DENMARK 48 125 ROMANIA ESTONIA 51 128 RUSSIA BirdLife Europe and Central Asia is a partnership of 48 national conservation organisations and a leader in bird conservation. Our unique local to global FAROE ISLANDS DENMARK 54 132 SERBIA approach enables us to deliver high impact and long term conservation for the beneit of nature and people. BirdLife Europe and Central Asia is one of FINLAND 56 135 SLOVAKIA the six regional secretariats that compose BirdLife International. Based in Brus- sels, it supports the European and Central Asian Partnership and is present FRANCE 60 138 SLOVENIA in 47 countries including all EU Member States. With more than 4,100 staf in Europe, two million members and tens of thousands of skilled volunteers, GEORGIA 64 141 SPAIN BirdLife Europe and Central Asia, together with its national partners, owns or manages more than 6,000 nature sites totaling 320,000 hectares. GERMANY 67 145 SWEDEN GIBRALTAR UNITED KINGDOM 71 148 SWITZERLAND GREECE 72 151 TURKEY GREENLAND DENMARK 76 155 UKRAINE HUNGARY 78 159 UNITED KINGDOM ICELAND 81 162 European population sizes and trends STICHTING BIRDLIFE EUROPE GRATEFULLY ACKNOWLEDGES FINANCIAL SUPPORT FROM THE EUROPEAN COMMISSION. -

Alpha Codes for 2168 Bird Species (And 113 Non-Species Taxa) in Accordance with the 62Nd AOU Supplement (2021), Sorted Taxonomically
Four-letter (English Name) and Six-letter (Scientific Name) Alpha Codes for 2168 Bird Species (and 113 Non-Species Taxa) in accordance with the 62nd AOU Supplement (2021), sorted taxonomically Prepared by Peter Pyle and David F. DeSante The Institute for Bird Populations www.birdpop.org ENGLISH NAME 4-LETTER CODE SCIENTIFIC NAME 6-LETTER CODE Highland Tinamou HITI Nothocercus bonapartei NOTBON Great Tinamou GRTI Tinamus major TINMAJ Little Tinamou LITI Crypturellus soui CRYSOU Thicket Tinamou THTI Crypturellus cinnamomeus CRYCIN Slaty-breasted Tinamou SBTI Crypturellus boucardi CRYBOU Choco Tinamou CHTI Crypturellus kerriae CRYKER White-faced Whistling-Duck WFWD Dendrocygna viduata DENVID Black-bellied Whistling-Duck BBWD Dendrocygna autumnalis DENAUT West Indian Whistling-Duck WIWD Dendrocygna arborea DENARB Fulvous Whistling-Duck FUWD Dendrocygna bicolor DENBIC Emperor Goose EMGO Anser canagicus ANSCAN Snow Goose SNGO Anser caerulescens ANSCAE + Lesser Snow Goose White-morph LSGW Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Intermediate-morph LSGI Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Blue-morph LSGB Anser caerulescens caerulescens ANSCCA + Greater Snow Goose White-morph GSGW Anser caerulescens atlantica ANSCAT + Greater Snow Goose Intermediate-morph GSGI Anser caerulescens atlantica ANSCAT + Greater Snow Goose Blue-morph GSGB Anser caerulescens atlantica ANSCAT + Snow X Ross's Goose Hybrid SRGH Anser caerulescens x rossii ANSCAR + Snow/Ross's Goose SRGO Anser caerulescens/rossii ANSCRO Ross's Goose -

The Breeding Population of Eurasian Dotterel <I>Charadrius Morinellus</I> in Spain
The breeding population of Eurasian Dotterel Charadrius morinellus in Spain Ricard Gutierrez,Antoni Carulla,Xavier Parellada,Diego Garcfa - FerrY, FrancescXavier Santaeufernia,Jordi Figuerola, Oriol Muntan & Francisco Cerd. Gutierrez,R., Carulla,A., Parellada,X., Garcia-FerrY,D., Santaeufemia,F.X., Figuerola,J., Muntan•, O. & Cerda, F. 1996. The breedingpopulation of Eurasian DotterelCharaddus morinellus•n Spain• Wader Study Group Bull. 80: 47-49. A partialcensus was carriedout in 1994 to confirmthe statusof EurasianDotterel in alpine areas of CatalonianPyrenees (NE Spain). Only one of the three areas, near the Andorranand Frenchfrontiers, gave positiveresults, with 2-4 pairs and two nests being found at altitudesof 2580 m and 2720 m. The minimumestimate for the wholeof the area was 2-15 pairs. The currentdistribution coincides with that knownpreviously from the XVlII centuryand is connectedwith that of Frenchand suspectedAndorran populations. The speciesbreeds on stony high plateauxwith some wet areas where they feed. Relief and thus, lack of suitable habitats,together with soiltype influencinglikelihood of floodingcould be factorsdetermining distribution. Gutierrez,R. & Santaeufemia,F.X., ReservesNaturals Delta de/Uobregat,Departament d,Agricultura,Ramaderia i Pesca, Gran •a de les Corts Catalanes, 612-614 2n, E-08007 Barcelona,Spain. Carulla,A., Ca La Madeta,Cam[ de Tall(•s/n, Tall(•,E-25721 Bellverde Cerdanya,Spain. Parellada, X., Garcia-FerrY, D., Servel de Protecci(•i Gesti(• de la Fauna, D.G. Medi Natural, Gran Via de les Corts Catalanes, 612-614 2n, E-08007 Barcelona,Spain. Figuerola,J. Departamentde BiologiaAnimal (Vertebrats), Universitatde Barcelona,Av. Diagonal,625, E-08028 Barcelona,Spain. Muntan•, 0o, JacintVerdaguer, 14 4rt 1-a, E-08902 L'Hospitaletde Uobregat,Spainø Cerd•, F., Av. -

Dotterel; What's in a Name?
Dotterel; What’s in a name? Wader Quest Article number 16: 19/01/2021 Rick and Elis Simpson Eurasian Dotterel © Vojtěch Kubelka SUPPORTING SHOREBIRD CONSERVATION Registered Charity (England and Wales) 1183748 © Wader Quest 2021. All rights reserved. 2 The Dotterel Charadrius morinellus; when I first heard the name as a youngster it was one of those, almost mythical, species which, as far as I could see, I had little or no chance of ever seeing. In those days even an outing to Norfolk from our house in the home counties, let alone a holiday to the Cairngorms, was still only a dream; my family could not afford a car. It was an inaccessible bird and the fact that I was unlikely to see one any time soon, added to the aura of mysticism surrounding the bird, and the wonderful name highlighted the specialness of this bird to me. I had no appreciation of why the bird was called by this unusual name, I thought it was rather quaint, cute even, but in fact, nothing could be further from the truth. Human beings have a habit of disappointing me and bursting my bubbles of naivety, especially Eurasian Dotterel © Jullian Bhalerao when it comes to birds and their conservation, and when I discovered why the Dotterel was thus named a thought occurred to me; what is it about we humans that makes us feel so damn superior to every other living thing on this planet? If an animal or bird doesn’t behave in a manner in which we can find admiration, courage, speed, agility, we end up belittling the creature; witness such expressions as dumb animals, bird brained, silly cow, filthy pig and so on. -

Arctic Species Trend Index: Migratory Birds Index
CAFF Assessment Series Report September 2015 Arctic Species Trend Index: Migratory Birds Index ARCTIC COUNCIL Acknowledgements CAFF Designated Agencies: • Norwegian Environment Agency, Trondheim, Norway • Environment Canada, Ottawa, Canada • Faroese Museum of Natural History, Tórshavn, Faroe Islands (Kingdom of Denmark) • Finnish Ministry of the Environment, Helsinki, Finland • Icelandic Institute of Natural History, Reykjavik, Iceland • Ministry of Foreign Affairs, Greenland • Russian Federation Ministry of Natural Resources, Moscow, Russia • Swedish Environmental Protection Agency, Stockholm, Sweden • United States Department of the Interior, Fish and Wildlife Service, Anchorage, Alaska CAFF Permanent Participant Organizations: • Aleut International Association (AIA) • Arctic Athabaskan Council (AAC) • Gwich’in Council International (GCI) • Inuit Circumpolar Council (ICC) • Russian Indigenous Peoples of the North (RAIPON) • Saami Council This publication should be cited as: Deinet, S., Zöckler, C., Jacoby, D., Tresize, E., Marconi, V., McRae, L., Svobods, M., & Barry, T. (2015). The Arctic Species Trend Index: Migratory Birds Index. Conservation of Arctic Flora and Fauna, Akureyri, Iceland. ISBN: 978-9935-431-44-8 Cover photo: Arctic tern. Photo: Mark Medcalf/Shutterstock.com Back cover: Red knot. Photo: USFWS/Flickr Design and layout: Courtney Price For more information please contact: CAFF International Secretariat Borgir, Nordurslod 600 Akureyri, Iceland Phone: +354 462-3350 Fax: +354 462-3390 Email: [email protected] Internet: www.caff.is This report was commissioned and funded by the Conservation of Arctic Flora and Fauna (CAFF), the Biodiversity Working Group of the Arctic Council. Additional funding was provided by WWF International, the Zoological Society of London (ZSL) and the Convention on Migratory Species (CMS). The views expressed in this report are the responsibility of the authors and do not necessarily reflect the views of the Arctic Council or its members. -

Avibase Page 1Of 15
Avibase Page 1of 15 Col Location Date Start time Duration Distance Avibase - Bird Checklists of the World 1 Country or region: Skåne 2 Number of species: 445 3 Number of endemics: 0 4 Number of breeding endemics: 0 5 Number of globally threatened species: 22 6 Number of extinct species: 0 7 Number of introduced species: 1 8 Date last reviewed: 2012-01-20 9 10 Recommended citation: Lepage, D. 2019. Checklist of the birds of Skåne. Avibase, the world bird database. Retrieved from .https://avibase.bsc- eoc.org/checklist.jsp?lang=EN®ion=sesn&list=clements&format=1 [19/03/2019]. Make your observations count! Submit your data to ebird. -
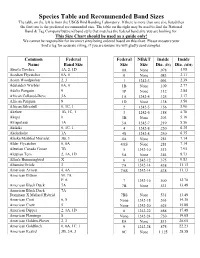
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American -

Family Anatidae Black-Bellied Whistling-Duck, 14 Fulvous Whistling-Duck, 14 Swan Goose, 3 Taiga Bean-Goose, 4 Tundra Bean-Go
Family Anatidae Hooded Merganser, 35 Flesh-footed Shearwater, 66 Cooper's Hawk, 98 Marbled Godwit, 135 Bonaparte's Gull, 157 Burrowing Owl, 217 American Three-toed Black-bellied Whistling-Duck, 14 Common Merganser, 34 Great Shearwater, 66 Northern Goshawk, 99 Ruddy Turnstone, 136 Black-headed Gull, 156 Spotted Owl, 215 Woodpecker, 252 Fulvous Whistling-Duck, 14 Red-breasted Merganser, 34 Buller's Shearwater, 64 Bald Eagle, 111 Black Turnstone, 136 Little Gull, 156 Barred Owl, 215 Black-backed Woodpecker, 252 Swan Goose, 3 Ruddy Duck, 35 Sooty Shearwater, 67 Common Black Hawk, 100 Great Knot, 137 Ross's Gull, 154 Great Gray Owl, 214 Northern Flicker, 253 Taiga Bean-Goose, 4 Family Numididae Short-tailed Shearwater, 67 Harris's Hawk, 100 Red Knot, 139 Laughing Gull, 159 Long-eared Owl, 212 Gilded Flicker, 253 Tundra Bean-Goose, 4 Helmeted Guineafowl, 50 Manx Shearwater, 65 Gray Hawk, 101 Surfbird, 139 Franklin's Gull, 158 Short-eared Owl, 212 Pileated Woodpecker, 242 Greater White-fronted Goose, 4 Family Odontophoridae Black-vented Shearwater, 65 Red-shouldered Hawk, 102 Ruff, 148 Black-tailed Gull, 174 Boreal Owl, 216 Ivory-billed Woodpecker, xvi Graylag Goose, 3 Mountain Quail, 39 Family Hydrobatidae Broad-winged Hawk, 103 Sharp-tailed Sandpiper, 140 Heermann's Gull, 173 Northern Saw-whet Owl, 216 Family Falconidae Bar-headed Goose, 3 Scaled Quail, 39 Wilson's Storm-Petrel, 68 Short-tailed Hawk, 105 Stilt Sandpiper, 146 Mew Gull, 160 Family Caprimulgidae Crested Caracara, 254 Emperor Goose, 8 California Quail, 38 Fork-tailed Storm-Petrel, -

Eudromias Morinellus
Eudromias morinellus -- (Linnaeus, 1758) ANIMALIA -- CHORDATA -- AVES -- CHARADRIIFORMES -- CHARADRIIDAE Common names: Eurasian Dotterel; Dotterel; Pluvier guignard European Red List Assessment European Red List Status LC -- Least Concern, (IUCN version 3.1) Assessment Information Year published: 2015 Date assessed: 2015-03-31 Assessor(s): BirdLife International Reviewer(s): Symes, A. Compiler(s): Ashpole, J., Burfield, I., Ieronymidou, C., Pople, R., Wheatley, H. & Wright, L. Assessment Rationale European regional assessment: Least Concern (LC) EU27 regional assessment: Least Concern (LC) In Europe this species has a very large range, and hence does not approach the thresholds for Vulnerable under the range size criterion (Extent of Occurrence 10% in ten years or three generations, or with a specified population structure). Despite the fact that the population trend appears to be decreasing, the decline is not believed to be sufficiently rapid to approach the thresholds for Vulnerable under the population trend criterion (30% decline over ten years or three generations). For these reasons the species is evaluated as Least Concern in Europe. Within the EU27 this species has a very large range, and hence does not approach the thresholds for Vulnerable under the range size criterion (Extent of Occurrence 10% in ten years or three generations, or with a specified population structure). Despite the fact that the population trend appears to be decreasing, the decline is not believed to be sufficiently rapid to approach the thresholds for Vulnerable under the population trend criterion (30% decline over ten years or three generations). For these reasons the species is evaluated as Least Concern in the EU27. Occurrence Countries/Territories of Occurrence Native: Albania; Andorra; Armenia; Austria; Belgium; Bulgaria; Cyprus; Czech Republic; Denmark; Estonia; Finland; France; Germany; Greece; Hungary; Ireland, Rep. -

American Ornithological Union (AOU) Bird Species List 1
American Ornithological Union (AOU) Bird Species List 1 Alpha Alpha Species Code Species Code Western Grebe WEGR Glaucous-winged Gull GWGU Clark's Grebe CLGR Hybrid Gull HYGU Red-necked Grebe RNGR Great Black-backed Gull GBBG Horned Grebe HOGR Slaty-backed Gull SBGU Eared Grebe EAGR Western Gull WEGU Least Grebe LEGR Yellow-footed Gull YFGU Pied-billed Grebe PBGR Lesser Black-backed Gull LBBG Common Loon COLO Herring Gull HERG Yellow-billed Loon YBLO California Gull CAGU Arctic Loon ARLO Unidentified Gull UNGU Pacific Loon PALO Ring-billed Gull RBGU Red-throated Loon RTLO Band-tailed Gull BTGU Tufted Puffin TUPU Mew Gull MEGU Atlantic Puffin ATPU Black-headed Gull BHGU Horned Puffin HOPU Heermann's Gull HEEG Rhinoceros Auklet RHAU Laughing Gull LAGU Cassin's Auklet CAAU Franklin's Gull FRGU Parakeet Auklet PAAU Bonaparte's Gull BOGU Crested Auklet CRAU Little Gull LIGU Whiskered Auklet WHAU Ross' Gull ROGU Least Auklet LEAU Sabine's Gull SAGU Ancient Murrelet ANMU Gull-billed Tern GBTE Marbled Murrelet MAMU Caspian Tern CATE Kittlitz's Murrelet KIMU Royal Tern ROYT Xantus' Murrelet XAMU Crested Tern CRTE Craveri's Murrelet CRMU Elegant Tern ELTE Black Guillemot BLGU Sandwich Tern SATE Pigeon Guillemot PIGU Cayenne Tern CAYT Common Murre COMU Forster's Tern FOTE Thick-billed Murre TBMU Common Tern COTE Razorbill RAZO Arctic Tern ARTE Dovekie DOVE Roseate Tern ROST Great Skua GRSK Aleutian Tern ALTE South Polar Skua SPSK Black-naped Tern BNTE Pomarine Jaeger POJA Least Tern LETE Parasitic Jaeger PAJA Sooty Tern SOTE Long-tailed Jaeger LTJA -

A Novel Approach to Detect Long-Term Changes in Terrestrial Faunal Abundance Using Historical Qualitative Descriptions
EUROPEAN JOURNAL OF ECOLOGY EJE 2015, 1(1): 32-42, doi: 10.1515/eje-2015-0005 Pushing back the baseline: a novel approach to detect long-term changes in terrestrial faunal abundance using historical qualitative descriptions 1 2 2,3,4,5 Duncan McCollin , Richard C. Preece , Tim H. Sparks 1Landscape and Biodi- ABSTRACT versity Research Group, Studies that examine changes in the populations of flora and fauna often do so against a baseline of relatively School of Science and recent distribution data. It is much rarer to see evaluations of population change over the longer–term in order Technology, The Univer- sity of Northampton, NN2 to extend the baseline back in time. Here, we use two methods (regression analysis and line of equality) to 6JD, UK, Corresponding identify long-term differences in abundance derived from qualitative descriptions, and we test the efficacy of Author, E-mail: Duncan. this approach by comparison with contemporary data. We take descriptions of bird population abundance in McCollin@northampton. Cambridgeshire, UK, from the first half of the 19th century and compare these with more recent estimates by ac.uk converting qualitative descriptions to an ordinal scale. We show, first, that the ordinal scale of abundance cor- 2Department of Zoology, responds well to quantitative estimates of density and range size based on current data, and, second, that the University of Cambridge, two methods of comparison revealed both increases and declines in species, some of which were consistent Cambridge CB2 3EJ, UK using both approaches but others showed differing responses. We also show that the regional rates of extinction 3Fachgebiet für Ökokli- (extirpation) for birds are twice as high as equivalent rates for plants.