Rodrigo Temp Müller
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Origin and Early Evolution of Dinosaurs
Biol. Rev. (2010), 85, pp. 55–110. 55 doi:10.1111/j.1469-185X.2009.00094.x The origin and early evolution of dinosaurs Max C. Langer1∗,MartinD.Ezcurra2, Jonathas S. Bittencourt1 and Fernando E. Novas2,3 1Departamento de Biologia, FFCLRP, Universidade de S˜ao Paulo; Av. Bandeirantes 3900, Ribeir˜ao Preto-SP, Brazil 2Laboratorio de Anatomia Comparada y Evoluci´on de los Vertebrados, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Avda. Angel Gallardo 470, Cdad. de Buenos Aires, Argentina 3CONICET (Consejo Nacional de Investigaciones Cient´ıficas y T´ecnicas); Avda. Rivadavia 1917 - Cdad. de Buenos Aires, Argentina (Received 28 November 2008; revised 09 July 2009; accepted 14 July 2009) ABSTRACT The oldest unequivocal records of Dinosauria were unearthed from Late Triassic rocks (approximately 230 Ma) accumulated over extensional rift basins in southwestern Pangea. The better known of these are Herrerasaurus ischigualastensis, Pisanosaurus mertii, Eoraptor lunensis,andPanphagia protos from the Ischigualasto Formation, Argentina, and Staurikosaurus pricei and Saturnalia tupiniquim from the Santa Maria Formation, Brazil. No uncontroversial dinosaur body fossils are known from older strata, but the Middle Triassic origin of the lineage may be inferred from both the footprint record and its sister-group relation to Ladinian basal dinosauromorphs. These include the typical Marasuchus lilloensis, more basal forms such as Lagerpeton and Dromomeron, as well as silesaurids: a possibly monophyletic group composed of Mid-Late Triassic forms that may represent immediate sister taxa to dinosaurs. The first phylogenetic definition to fit the current understanding of Dinosauria as a node-based taxon solely composed of mutually exclusive Saurischia and Ornithischia was given as ‘‘all descendants of the most recent common ancestor of birds and Triceratops’’. -

The Anatomy and Phylogenetic Relationships of Antetonitrus Ingenipes (Sauropodiformes, Dinosauria): Implications for the Origins of Sauropoda
THE ANATOMY AND PHYLOGENETIC RELATIONSHIPS OF ANTETONITRUS INGENIPES (SAUROPODIFORMES, DINOSAURIA): IMPLICATIONS FOR THE ORIGINS OF SAUROPODA Blair McPhee A dissertation submitted to the Faculty of Science, University of the Witwatersrand, in partial fulfilment of the requirements for the degree of Master of Science. Johannesburg, 2013 i ii ABSTRACT A thorough description and cladistic analysis of the Antetonitrus ingenipes type material sheds further light on the stepwise acquisition of sauropodan traits just prior to the Triassic/Jurassic boundary. Although the forelimb of Antetonitrus and other closely related sauropododomorph taxa retains the plesiomorphic morphology typical of a mobile grasping structure, the changes in the weight-bearing dynamics of both the musculature and the architecture of the hindlimb document the progressive shift towards a sauropodan form of graviportal locomotion. Nonetheless, the presence of hypertrophied muscle attachment sites in Antetonitrus suggests the retention of an intermediary form of facultative bipedality. The term Sauropodiformes is adopted here and given a novel definition intended to capture those transitional sauropodomorph taxa occupying a contiguous position on the pectinate line towards Sauropoda. The early record of sauropod diversification and evolution is re- examined in light of the paraphyletic consensus that has emerged regarding the ‘Prosauropoda’ in recent years. iii ACKNOWLEDGEMENTS First, I would like to express sincere gratitude to Adam Yates for providing me with the opportunity to do ‘real’ palaeontology, and also for gladly sharing his considerable knowledge on sauropodomorph osteology and phylogenetics. This project would not have been possible without the continued (and continual) support (both emotionally and financially) of my parents, Alf and Glenda McPhee – Thank you. -

Ontogenetic Changes in the Body Plan of the Sauropodomorph Dinosaur Mussaurus Patagonicus Reveal Shifts of Locomotor Stance Duri
www.nature.com/scientificreports OPEN Ontogenetic changes in the body plan of the sauropodomorph dinosaur Mussaurus patagonicus Received: 8 November 2018 Accepted: 8 May 2019 reveal shifts of locomotor stance Published: xx xx xxxx during growth Alejandro Otero 1,2, Andrew R. Cuf 3, Vivian Allen3, Lauren Sumner-Rooney3,4, Diego Pol2,5 & John R. Hutchinson 3 Ontogenetic information is crucial to understand life histories and represents a true challenge in dinosaurs due to the scarcity of growth series available. Mussaurus patagonicus was a sauropodomorph dinosaur close to the origin of Sauropoda known from hatchling, juvenile and mature specimens, providing a sufciently complete ontogenetic series to reconstruct general patterns of ontogeny. Here, in order to quantify how body shape and its relationship with locomotor stance (quadruped/ biped) changed in ontogeny, hatchling, juvenile (~1 year old) and adult (8+ years old) individuals were studied using digital models. Our results show that Mussaurus rapidly grew from about 60 g at hatching to ~7 kg at one year old, reaching >1000 kg at adulthood. During this time, the body’s centre of mass moved from a position in the mid-thorax to a more caudal position nearer to the pelvis. We infer that these changes of body shape and centre of mass refect a shift from quadrupedalism to bipedalism occurred early in ontogeny in Mussaurus. Our study indicates that relative development of the tail and neck was more infuential in determining the locomotor stance in Sauropodomorpha during ontogeny, challenging previous studies, which have emphasized the infuence of hindlimb vs. forelimb lengths on sauropodomorph stance. -

Skull Remains of the Dinosaur Saturnalia Tupiniquim (Late Triassic, Brazil): with Comments on the Early Evolution of Sauropodomorph Feeding Behaviour
RESEARCH ARTICLE Skull remains of the dinosaur Saturnalia tupiniquim (Late Triassic, Brazil): With comments on the early evolution of sauropodomorph feeding behaviour 1 2 1 Mario BronzatiID *, Rodrigo T. MuÈ llerID , Max C. Langer * 1 LaboratoÂrio de Paleontologia, Faculdade de Filosofia Ciências e Letras de Ribeirão Preto, Universidade de a1111111111 São Paulo, Ribeirão Preto, São Paulo, Brazil, 2 Centro de Apoio à Pesquisa PaleontoloÂgica, Universidade Federal de Santa Maria, Santa Maria, Rio Grande do Sul, Brazil a1111111111 a1111111111 * [email protected] (MB); [email protected] (MCL) a1111111111 a1111111111 Abstract Saturnalia tupiniquim is a sauropodomorph dinosaur from the Late Triassic (Carnian±c. 233 OPEN ACCESS Ma) Santa Maria Formation of Brazil. Due to its phylogenetic position and age, it is important for studies focusing on the early evolution of both dinosaurs and sauropodomorphs. The Citation: Bronzati M, MuÈller RT, Langer MC (2019) Skull remains of the dinosaur Saturnalia tupiniquim osteology of Saturnalia has been described in a series of papers, but its cranial anatomy (Late Triassic, Brazil): With comments on the early remains mostly unknown. Here, we describe the skull bones of one of its paratypes (only in evolution of sauropodomorph feeding behaviour. the type-series to possess such remains) based on CT Scan data. The newly described ele- PLoS ONE 14(9): e0221387. https://doi.org/ ments allowed estimating the cranial length of Saturnalia and provide additional support for 10.1371/journal.pone.0221387 the presence of a reduced skull (i.e. two thirds of the femoral length) in this taxon, as typical Editor: JuÈrgen Kriwet, University of Vienna, of later sauropodomorphs. -

Terræ Evidências Da Origem E Ascensão Dos Dinossauros
Terræ ARTIGO 10.20396/td.v16i0.8657367 Didatica Evidências da origem e ascensão dos dinossauros sauropodomorfos preservadas em leitos fossilíferos do Triássico do Sul do Brasil EVIDENCE OF THE ORIGIN AND RISE OF SAUROPODOMORPH DINOSAURS PRESERVED IN TRIASSIC BEDS FROM SOUTHERN BRAZIL RODRIGO TEMP MÜLLER1, MAURÍCIO SILVA GARCIA2, SÉRGIO DIAS-DA-SILVA2 1 - CENTRO APOIO PESQ. PALEONT. DA QUARTA COLÔNIA, UNIV. FED. SANTA MARIA, SÃO JOÃO DO POLÊSINE, BRASIL. 2 - LABORATÓRIO DE PALEOBIODIVERSIDADE TRIÁSSICA, DEPARTAMENTO DE ECOLOGIA E EVOLUÇÃO, UNIV. FED. SANTA MARIA, SANTA MARIA, BRASIL. E-MAIL: [email protected], [email protected], [email protected] Abstract: Although the sauropodomorph dinosaur group is widely known due to the giant sauropods, Citation/Citação: Müller, R. T., Garcia, their origin is still poorly understood due to the scarcity of complete skeletons and precise dating. M. S., & Silva, S. D. (2020). Evidências However, new findings and dating performed in Triassic rocks from Brazil have brought forth valuable da origem e ascensão dos dinossauros information regarding this matter. The combination of new data and more advanced investigation sauropodomorfos preservadas em leitos techniques (e.g. CT scans) have enabled the establishment of macroevolutionary patterns that guided fossilíferos do Triássico do Sul do Brasil. the initial evolution of the group. Through discoveries made in Triassic beds of Rio Grande do Sul Terræ Didatica, 16, 1-13, e020013. state, we can understand in more detail how sauropodomorphs went from small and not abundant doi:10.20396/td.v16i0.8657367 animals to the largest animals that ever walked the Earth. Keywords: Dinosauria. Evolution. Mesozoic Resumo: Embora o grupo dos dinossauros sauropodomorfos seja amplamente conhecido em virtude Era. -

The First Dinosaur Egg Remains a Mystery
bioRxiv preprint doi: https://doi.org/10.1101/2020.12.10.406678; this version posted December 11, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 The first dinosaur egg remains a mystery 2 3 Lucas J. Legendre1*, David Rubilar-Rogers2, Alexander O. Vargas3, and Julia A. 4 Clarke1* 5 6 1Department of Geological Sciences, University of Texas at Austin, Austin, Texas 78756, 7 USA. 8 2Área Paleontología, Museo Nacional de Historia Natural, Casilla 787, Santiago, Chile. 9 3Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Santiago 7800003, 10 Chile. 11 1 bioRxiv preprint doi: https://doi.org/10.1101/2020.12.10.406678; this version posted December 11, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 12 Abstract 13 A recent study by Norell et al. (2020) described new egg specimens for two dinosaur species, 14 identified as the first soft-shelled dinosaur eggs. The authors used phylogenetic comparative 15 methods to reconstruct eggshell type in a sample of reptiles, and identified the eggs of 16 dinosaurs and archosaurs as ancestrally soft-shelled, with three independent acquisitions of a 17 hard eggshell among dinosaurs. This result contradicts previous hypotheses of hard-shelled 18 eggs as ancestral to archosaurs and dinosaurs. -

Dinosaurs.Pdf
DINOSAURS COMPILED BY HOWIE BAUM THIS IS THE FIRST OF 3 PAGES THAT HELP TO PUT THE 165 MILLION YEAR REIGN OF THE DINOSAURS, IN PERSPECTIVE. IT IS A CALENDAR THAT IS DESIGNED SO THAT ALL OF THE TIME, SINCE THE BIG BANG HAPPENED – 13.8 BILLION YEAR AGO, UP TO THE PRESENT TIME, IS COMPRESSED INTO 1 YEAR. period First Dinosaurs Dinosaurs lived for 165 million years !! Humans have only existed for 0.004% of the age of the Earth ! The rocks of the Cincinnatian series were deposited between approximately 451 and 443 million years ago, during the Ordovician period, when all the land was underwater ! BASIC DINOSAUR FACTS •Dinosaurs are a group of reptiles that have lived on Earth for about 165 million years. About 60% of dinosaurs ate plants (herbivores) and 40% ate meat (carnivores) •In 1842, the English naturalist Sir Richard Owen coined the term Dinosauria, derived from the Greek deinos, meaning “fearfully great,” and sauros, meaning “lizard.” •Dinosaur fossils have been found on all 7 continents. •All non-avian (non-bird) dinosaurs went extinct about 66 million years ago (MYA). •There are roughly 700 known species of extinct dinosaurs. •Modern birds are a kind of dinosaur because they share a common ancestor with non-avian dinosaurs. The Archosaurs consist of a diverse group of Triassic living and extinct reptiles that are subdivided into crocodiles, the 2 main types of dinosaurs, and pterosaurs. ORNITHISCHIAN AND SAURISCHIAN DINOSAURS As shown on the previous diagram, there are 2 types of Dinosaurs. Dinosaurs and reptiles have hip girdles, or pelvises, and all of them are composed of three bones: the ilium, ischium, and pubis. -

What Are Dinosaurs?
Tote Hughes, 140819 [email protected] Dinosaurs 1/20 What Are Dinosaurs? The following are not dinosaurs∗: • Things that aren’t organisms—There is no rock that is a dinosaur. • Things that existed before the Triassic period† • Pterosaurs The following are dinosaurs: • Birds (Aves) The following contain dinosaurs: • Archosaurs (X.Archosauria) • Reptiles (Reptilia) Dinosaur Overview A discussion of the important dinosaur clades. Dinosaurs are divided into two main groups: the eusaurischians‡ and ornithischians. Eusaurischians • Sauropods – Apatasaurus: diplodocoidean – Barosaurus: diplodocoidean – Brachiosaurus: macronarian – Diplodocus: diplodocoidean • Theropods – Allosaurus: carnosaurian – Archaeopteryx: maniraptor – Giganotosaurus: carnosaurian – Megalosaurus: megalosaurid – Spinosaurus: megalosaurid – Tyrannosaurus: tyrannosauroid – Velociraptor: maniraptor ∗See the Dinosaur Encyclopedia section for details on terms. †See Appendix: Time for details on geological time. ‡These are commonly called saurischians, but since almost every interesting saurischian is actually in the subclade X.Eusaurischia, I’ve taken the liberty of breaking the standard. I hope you will grow to understand and accept my decision. 1/20 Tote Hughes, 140819 [email protected] Dinosaurs 2/20 Ornithischians • Eurypodans (thyreophor) – Ankylosaurus: ankylosaurian – Stegosaurus: stegosaurian • Marginocephalians (cerapod) – Pachycephalosaurus: pachycephalosaurian – Triceratops: ceratopsian • Ornithopods (cerapod) – Hadrosaurus: hadrosauriform – Iguanodon: hadrosauriform -
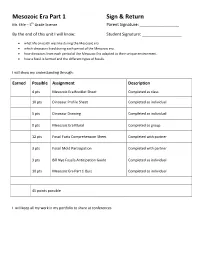
Mesozoic Era Part 1 Sign & Return
Mesozoic Era Part 1 Sign & Return Mr. Ehle – 5 th Grade Science Parent Signature: ______________________ By the end of this unit I will know: Student Signature: ______________________ • what life on earth was like during the Mesozoic era • which dinosaurs lived during each period of the Mesozoic era. • how dinosaurs from each period of the Mesozoic Era adapted to their unique environment. • how a fossil is formed and the different types of fossils. I will show my understanding through: Earned Possible Assignment Description 4 pts Mesozoic Era Booklet Sheet Completed as class 10 pts Dinosaur Profile Sheet Completed as individual 5 pts Dinosaur Drawing Completed as individual 0 pts Mesozoic Era Mural Completed as group 12 pts Fossil Facts Comprehension Sheet Completed with partner 3 pts Fossil Mold Participation Completed with partner 3 pts Bill Nye Fossils Anticipation Guide Completed as individual 10 pts Mesozoic Era Part 1 Quiz Completed as individual 45 points possible I will keep all my work in my portfolio to share at conferences. Mesozoic Era Part 1 Study Guide Vocabulary Mesozoic Era An era of geologic time that lasted from 245-65 million years ago Triassic Period 245-208 Million Years Ago Jurassic Period 208-135 Million Years Ago Cretaceous Period 135-65 Million Years Ago adapt To adjust to particular conditions extinction The total disappearance of a species so that it no longer exists amber tree sap that became hard fossil bones and other signs of a plant or animal paleontologist scientist who studies fossils trace fossil fossil that shows just a sign of something, like a footprint The Mesozoic Era is so long that it is divided into 3 parts: Triassic, Jurassic and Cretaceous periods. -

El Registro De Sauropodomorpha (Dinosauria) De La Argentina
ISSN 2469-0228 www.peapaleontologica.org.ar EL REGISTRO DE SAUROPODOMORPHA (DINOSAURIA) DE LA ARGENTINA ALEJANDRO OTERO 1 LEONARDO SALGADO 2 1CONICET. División Paleontología Vertebrados, Museo de La Plata. Paseo del Bosque s/n, B1900FWA La Plata, Argentina. 2CONICET. Instituto de Investigación en Paleobiología y Geología, Universidad Nacional de Río Negro. Av. General Roca 1242, R8332EXZ General Roca, Río Negro, Argentina. Recibido: 4 de Junio de 2015 - Aceptado: 12 de Agosto de 2015 Para citar este artículo: Alejandro Otero y Leonardo Salgado (2015). El registro de Sauropodomorpha (Dinosauria) de la Argentina. En: M. Fernández y Y. Herrera (Eds.) Reptiles Extintos - Volumen en Homenaje a Zulma Gasparini . Publicación Electrónica de la Asociación Paleontológica Argentina 15(1): 69–89. Link a este artículo: http://dx.doi.org/ 10.5710/PEAPA.04.06.2015.100 DESPLAZARSE HACIA ABAJO PARA ACCEDER AL ARTÍCULO Asociación Paleontológica Argentina Maipú 645 1º piso, C1006ACG, Buenos Aires República Argentina Tel/Fax (54-11) 4326-7563 Web: www.apaleontologica.org.ar Otros artículos en Publicación Electrónica de la APA 15(1): de la Fuente & Sterli Paulina Carabajal Pol & Leardi ESTADO DEL CONOCIMIENTO DE GUIA PARA EL ESTUDIO DE LA DIVERSITY PATTERNS OF LAS TORTUGAS EXTINTAS DEL NEUROANATOMÍA DE DINOSAURIOS NOTOSUCHIA (CROCODYLIFORMES, TERRITORIO ARGENTINO: UNA SAURISCHIA, CON ENFASIS EN MESOEUCROCODYLIA) DURING PERSPECTIVA HISTÓRICA. FORMAS SUDAMERICANAS. THE CRETACEOUS OF GONDWANA. Año 2015 - Volumen 15(1): 69-89 VOLUMEN TEMÁTICO ISSN 2469-0228 EL REGISTRO DE SAUROPODOMORPHA (DINOSAURIA) DE LA ARGENTINA ALEJANDRO OTERO 1 Y LEONARDO SALGADO 2 1CONICET. División Paleontología Vertebrados, Museo de La Plata. Paseo del Bosque s/n, B1900FWA La Plata, Argentina. -

Cranial Ontogenetic Variation in Early Saurischians and the Role of Heterochrony in the Diversification of Predatory Dinosaurs
A peer-reviewed version of this preprint was published in PeerJ on 18 January 2016. View the peer-reviewed version (peerj.com/articles/1589), which is the preferred citable publication unless you specifically need to cite this preprint. Foth C, Hedrick BP, Ezcurra MD. 2016. Cranial ontogenetic variation in early saurischians and the role of heterochrony in the diversification of predatory dinosaurs. PeerJ 4:e1589 https://doi.org/10.7717/peerj.1589 Cranial ontogenetic variation in early saurischians and the role of heterochrony in the diversification of predatory dinosaurs Foth Christian, Brandon P Hedrick, Martin D Ezcurra Non-avian saurischian skulls underwent at least 165 million years of evolution and shapes varied from elongated skulls, such as in Coelophysis, to short and box-shaped skulls, such as in Camarasaurus. A number of factors have long been considered to drive skull shape, including phylogeny, dietary preferences and functional constraints. However, heterochrony is increasingly being recognized as a major factor in dinosaur evolution. In order to quantitatively analyse the impact of heterochrony on saurischian skull shape, we have analysed five ontogenetic trajectories using two-dimensional geometric PrePrints morphometrics in a phylogenetic framework. This allowed for the evaluation of how heterochrony affected overall skull shape through both ontogenetic and phylogenetic trajectories and how it impacted modular changes within the skull. Using principal component analyses and multivariate regressions, it was possible to quantify different ontogenetic trajectories in light of heterochrony. The results recovered here indicate that taxa underwent a combination of local paedomorphosis and peramorphosis within the skull along individual ontogenies and phylogenies, but that either peramorphosis or paedomorphosis dominated when the skull was considered as a whole. -

The Postcranial Anatomy of Coloradisaurus Brevis (Dinosauria
[Palaeontology, 2012, pp. 1–25] THE POSTCRANIAL ANATOMY OF COLORADISAURUS BREVIS (DINOSAURIA: SAUROPODOMORPHA) FROM THE LATE TRIASSIC OF ARGENTINA AND ITS PHYLOGENETIC IMPLICATIONS by CECILIA APALDETTI1*, DIEGO POL2 and ADAM YATES3 1CONICET, Universidad Nacional de San Juan, Museo de Ciencias Naturales, Av. Espana 400 (norte), San Juan, 5400 Argentina; e-mail: [email protected] 2CONICET, Museo Paleontologico Egidio Feruglio, Av. Fontana 140, Trelew, Chubut 9100, Argentina; e-mail: [email protected] 3University of the Witwatersrand, Bernard Price Institute for Palaeontological Research, Private Bag 3, Johannesburg, Gauteng, 2050, South Africa; e-mail: [email protected] *Corresponding author. Typescript received February 2012; accepted in revised form 13 July 2012 Abstract: Basal sauropodomorphs from the Upper Triassic surface of the tibia deflected and facing posterodistally and Los Colorados Formation of north-western Argentina have well-developed pyramidal dorsal process of the posteromedial been known for several decades but most of them are only corner of the astragalus. Several postcranial characters of briefly described. New postrcanial remains of Coloradisaurus Coloradisaurus are exclusively shared with Lufengosaurus, brevis, the most gracile sauropodomorph from this unit, are from the Lower Jurassic of China. The inclusion of this described here and evaluated within a phylogenetic context. information in two recent phylogenetic data sets depicts These materials belong to a single individual and include ele- Coloradisaurus as closely related to Lufengosaurus and well ments of the vertebral column, pectoral girdle, incomplete nested within Plateosauria. Both data sets used indicate forelimb, pelvis and hindlimb. These elements share an auta- strong character support for the inclusion of Coloradisaurus pomorphic feature with the type specimen of Coloradisaurus and Lufengosaurus within Massospondylidae.