Palynology of Badger Coprolites from Central Spain
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Wood Cutting Permit
CITY OF CRETE DEPARTMENTS OF PUBLIC WORKS • Permit to enter City property for purposes of woodcutting and wood hauling Conditions: 1. A valid permit is required to enter City property for purposes of cutting and hauling wood. 2. Permit is valid only for the undersigned as listed. 3. Permit only valid during the hours between 8:00 am to 8:00 pm Monday through Saturday and 10:00 am to 8:00 pm on Sunday. 4. City personnel will not aid or assist in cutting or loading. 5. Each permit holder is responsible for their own personal safety. The City of Crete assumes no responsibility or liability for permit holder or accompanying parties in case of accidents. 6. Permit holders are expected to be courteous of others. The City reserves the right to expel permit holders for any reason. 7. This wood is for individual personal use only. No resale for profit or commercial use is allowed. 8. By signing below, applicant agrees to all terms as listed above. Applicant will also read and sign the release statement accompanying this permit for permit to be valid. 9. This permit is valid from January 1 to December 31. 10.$10.00 fee charged to residents outside of City Limits. Name: _____________________________________________________________________ License number of vehicle/and trailer to be used: ___________________________________ Date permit issued: ___________________________________________________________ Signature of applicant: _________________________________________________________ Issued by: _____ City of Crete ! Departments of Public Works ! 243 -

Hand-Arm Vibration in the Aetiology of Hearing Loss in Lumberjacks
British Journal of Industrial Medicine 1981 ;38 :281-289 Hand-arm vibration in the aetiology of hearing loss in lumberjacks I PYYKKO,l J STARCK,2 M FARKKILA,l M HOIKKALA,2 0 KORHONEN,2 AND M NURMINEN2 From the Institute ofPhysiology,' University of Helsinki, and the Institute of Occupational Health,2 Helsinki, Finland ABSTRACT A longitudinal study of hearing loss was conducted among a group of lumberjacks in the years 1972 and 1974-8. The number of subjects increased from 72 in 1972 to 203 in 1978. They were classified according to (1) a history of vibration-induced white finger (VWF), (2) age, (3) duration of exposure, and (4) duration of ear muff usage. The hearing level at 4000 Hz was used to indicate the noise-induced permanent threshold shift (NIPTS). The lumberjacks were exposed, at their present pace of work, to noise, Leq values 96-103 dB(A), and to the vibration of a chain saw (linear acceleration 30-70 ms-2). The chain saws of the early 1960s were more hazardous, with the average noise level of 111 dB(A) and a variation acceleration of 60-180 ms2. When classified on the basis of age, the lumberjacks with VWF had about a 10 dB greater NIPTS than subjects without VWF. NIPTS increased with the duration of exposure to chain saw noise, but with equal noise exposure the NIPTS was about 10 dB greater in lumberjacks with VWF than without VWF. With the same duration of ear protection the lumberjacks with VWF consistently had about a 10 dB greater NIPTS than those without VWF. -

Kelly Mantle
The VARIETY SHOW With Your Host KELLY MANTLE KELLY MANTLE can be seen in the feature film Confessions of a Womanizer, for which they made Oscars history by being the first person ever to be approved and considered by The Academy for both Supporting Actor and Supporting Actress. This makes Kelly the first openly non-binary person to be considered for an Oscar. They are also featured in the movie Middle Man and just wrapped production on the upcoming feature film, God Save The Queens in which Kelly is the lead in. TV: Guest-starred on numerous shows, including Lucifer, Modern Family, Curb Your Enthusiasm, CSI, The New Normal, New Adventures of Old Christine, Judging Amy, Nip/Tuck, Will & Grace, George Lopez. Recurring: NYPD Blue. Featured in LOGO’s comedy special DragTastic NYC, and a very small co-star role on Season Six of RuPaul's Drag Race. Stage: Kelly has starred in more than 50 plays. They wrote and starred in their critically acclaimed solo show,The Confusion of My Illusion, at the Los Angeles LGBT Center. As a singer, songwriter, and musician, Kelly has released four critically acclaimed albums and is currently working on their fourth. Kelly grew up in Oklahoma like their uncle, the late great Mickey Mantle. (Yep...Kelly's a switch-hitter, too.) Kelly received a B.F.A. in Theatre from the University of Oklahoma and is a graduate of Second City in Chicago. https://www.instagram.com/kellymantle • https://www.imdb.com/name/nm0544141/ ALEXANDRA BILLINGS is an actress, teacher, singer, and activist. -

Virginia Commonwealth University Volunteer Doula Program Training Manual Kathleen M
Virginia Commonwealth University VCU Scholars Compass School of Nursing Publications School of Nursing 2015 Virginia Commonwealth University Volunteer Doula Program Training Manual Kathleen M. Bell Virginia Commonwealth University, [email protected] Susan L. Linder Virginia Commonwealth University, [email protected] Follow this and additional works at: http://scholarscompass.vcu.edu/nursing_pubs Part of the Maternal, Child Health and Neonatal Nursing Commons Copyright © 2015 The Authors Downloaded from http://scholarscompass.vcu.edu/nursing_pubs/16 This Curriculum Material is brought to you for free and open access by the School of Nursing at VCU Scholars Compass. It has been accepted for inclusion in School of Nursing Publications by an authorized administrator of VCU Scholars Compass. For more information, please contact [email protected]. Virginia Commonwealth University Volunteer Doula Program Training Manual “Empowerment, advocacy and support for one of life’s greatest journeys.” 1 Reflection and Discussion Why are you here? Why did you decide to do this doula training? What experiences do you have with birth, and how have they shaped your desire to participate in this program? What does it mean to be a doula with the VCU School of Nursing? What other reflections do you have? 2 Table of Contents 1. An overview of birth: Statistics and trends………………page 4-6 2. Birth workers and their roles………………...……………pages 7-10 3. You are a doula! Your birth-bag and preparation………..pages 11-14 4. Anatomy and Physiology of birth………………………...pages 15-18 5. Hormonal regulation of labor and birth…………………..pages 19-21 6. Pharmacologic management of labor…………………….pages 22-26 7. -

Comparisons of Laterality Between Wolves and Domesticated Dogs (Canis Lupus Familiaris)
Comparisons of laterality between wolves and domesticated dogs (Canis lupus familiaris) By Lindsey Drew Submitted to the Board of Biology School of Natural Sciences in partial fulfillment of the requirements for the degree of Bachelor of Science Purchase College State University of New York May 2015 ______________________________ Sponsor: Dr. Lee Ehrman _______________________________ Co-Sponsor: Rebecca Bose of the Wolf Conservation Center Table of Contents Abstract Page 2 Introduction Pages 3-18 Materials and Methods Pages 18-25 Results Pages 25-28 Discussion Pages 29-30 1 Abstract I compared laterality in Canis lupus familiaris (domesticated dog) with: Canis lupus occidentalis (Canadian Rocky Mountain wolves), Canis rufus (red wolves), Canis lupus baileyi (Mexican wolves), and Canis lupus arctos (Artic wolf), all related. Each wolf species was grouped into a single category and was compared to domesticated dogs. The methods performed on domesticated dogs were the KongTM test and the step test. For wolves the PVC test was created to simulate the KongTM test and a step test for comparison. My hypothesis was that Canis lupus familiaris and Canis lupus will show consistent lateralized preferences between both species. The significance of this study is that laterality in Canis lupus familiaris has been studied multiple times with varying results. The purpose is to determine the possibility of laterality in Canis lupus familiaris being a conserved trait. This study has the potential to give greater meaning to laterality and the benefits of right or left sidedness or handedness. The total amount of paw touches recorded under each category, for the step test in dogs were 280 (50.5%) left and 274 (49.5%) right, for the KongTM test there were 278 left (50.3%) and 275 right (49.7%). -

New Season of Hgtv's Hit Series 'Good Bones
NEW SEASON OF HGTV’S HIT SERIES ‘GOOD BONES’ SPOTLIGHTS DRAMATIC HOME RENOVATIONS AND FOLLOWS MINA STARSIAK HAWK’S IVF JOURNEY New Episodes Premiere Tuesday, June 9, at 9 p.m. ET/PT New York [May 12, 2020] Crumbling roofs, bug-infested walls and rotted floors are no match for HGTV’s popular mother/daughter duo Mina Starsiak Hawk and Karen E Laine in the new season of HGTV’s Good Bones, premiering Tuesday, June 9, at 9 p.m. ET/PT. In the series, which attracted more than 17.4 million total viewers in its last season, cameras follow Mina, a real estate agent and soon-to-be mom of two, and Karen, a lawyer, as they buy the most dilapidated properties in their favorite Indianapolis neighborhoods, demo them down to the studs and completely transform them into gorgeous, functional family homes. This season also spotlights very personal moments for both mother and daughter—Mina shares her emotional IVF journey that results in her second pregnancy while Karen announces she is retiring from the day-to-day operations of the family renovation business. “It’s been a very busy year renovating homes, opening our new storefront, navigating Mom’s retirement and dealing with the roller coaster of trying to get pregnant, all while raising a toddler,” said Mina. “I’m excited to share every step with our fans.” “I decided it was the right time to retire from our business, but I’ll still be renovating homes with Mina in the city we love,” said Karen. “The properties in this season are some of the best we’ve ever found.” In the premiere episode, Mina and Karen buy their most expensive house to date—a bungalow in the trendy Fountain Square neighborhood that needs serious updates. -

Coywolf: Eastern Coyote Genetics, Ecology, Management, and Politics
Coywolf: Eastern Coyote Genetics, Ecology, Management, and Politics By Jonathan G. Way Published by Eastern Coyote/Coywolf Research - www.EasternCoyoteResearch.com E-book • Citation: • Way, J.G. 2021. E-book. Coywolf: Eastern Coyote Genetics, Ecology, Management, and Politics. Eastern Coyote/Coywolf Research, Barnstable, Massachusetts. 277 pages. Open Access URL: http://www.easterncoyoteresearch.com/CoywolfBook. • Copyright © 2021 by Jonathan G. Way, Ph.D., Founder of Eastern Coyote/Coywolf Research. • Photography by Jonathan Way unless noted otherwise. • All rights reserved. No part of this book may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, e-mailing, or by any information storage, retrieval, or sharing system, without permission in writing or email to the publisher (Jonathan Way, Eastern Coyote Research). • To order a copy of my books, pictures, and to donate to my research please visit: • http://www.easterncoyoteresearch.com/store or MyYellowstoneExperience.org • Previous books by Jonathan Way: • Way, J. G. 2007 (2014, revised edition). Suburban Howls: Tracking the Eastern Coyote in Urban Massachusetts. Dog Ear Publishing, Indianapolis, Indiana, USA. 340 pages. • Way, J. G. 2013. My Yellowstone Experience: A Photographic and Informative Journey to a Week in the Great Park. Eastern Coyote Research, Cape Cod, Massachusetts. 152 pages. URL: http://www.myyellowstoneexperience.org/bookproject/ • Way, J. G. 2020. E-book (Revised, 2021). Northeastern U.S. National Parks: What Is and What Could Be. Eastern Coyote/Coywolf Research, Barnstable, Massachusetts. 312 pages. Open Access URL: http://www.easterncoyoteresearch.com/NortheasternUSNationalParks/ • Way, J.G. 2020. E-book (Revised, 2021). The Trip of a Lifetime: A Pictorial Diary of My Journey Out West. -
The Bones Short List
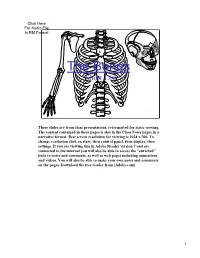
The Bones Short List 1 These slides are from class presentations, reformatted for static viewing. The content contained in these pages is also in the Class Notes pages in a narrative format. Best screen resolution for viewing is 1024 x 768. To change resolution click on start, then control panel, then display, then settings. If you are viewing this in Adobe Reader version 7 and are connected to the internet you will also be able to access the “enriched” links to notes and comments, as well as web pages including animations and videos. You will also be able to make your own notes and comments on the pages. Download the free reader from [Adobe.com] 1 Types of Bones 1) Long bones – bones of the upper and lower arms and legs as well as the metacarpals and metatarsals. 2) flat bones – sternum, ribs, scapulae, cranium 3) short bones – carpals and tarsals 4) irregular bones – vertebrae 5) sesamoid bones – patellae 2 2 Skeletal Divisions 1) Axial skeleton – skull, sternum, ribs, and vertebrae (including sacrum) 2) Appendicular skeleton – bones of the pectoral and pelvic girdles and of the arms, legs, wrists, hands, ankles, and feet. 3 3 Figure 7.3 a Frontal (coronal) suture Sphenoid Parietal bone bone Squamosal Frontal bone (temporal) suture Occipital (lamdoidal) Nasal bone suture Zygomatic bone Occipital bone Maxilla Temporal bone Mandible 4 Important: the mastoid process (important site for attachment of muscles such as the sternocleidomastoid), the external acoustic meatus (opening into the parts of the ear), the articulation of the temporal bone and mandible ( the TMJ), and the zygomatic arch, all on the temporal bone. -

Download Bare Bones, Kathy Reichs, Random House, 1999
Bare Bones, Kathy Reichs, Random House, 1999, , . DOWNLOAD http://archbd.net/17PVC8q Bones are Forever , Kathy Reichs, 2013, Fiction, 414 pages. The gripping new Temperance Brennan novel from the world class forensic anthropologist and Number 1 bestselling author.A newborn baby is found wedged in a vanity cabinet in a .... The Law of Betrayal , Tess Collins, Jun 30, 2006, Fiction, 292 pages. When she was ten years old Alma's father disappeared. The only man who knows the true story is brutally killed, but she must defend herself against an accusation of murder.. Break No Bones A Novel, Kathy Reichs, Jul 11, 2006, Fiction, 352 pages. Following the tremendous success of Cross Bones, Kathy Reichs explores another high-profile topic in Break No Bones -- a case that lands forensic anthropologist Temperance .... Bones Buried Deep, Max Allan Collins, Kathy Reichs, Feb 28, 2006, Fiction, 304 pages. Forensic anthropologist Dr. Termperance Brennan is called in by Special Agent Seeley Booth to assist in the investigation into a bag of skeletal remains, complete with note .... Cross Bones , Kathy Reichs, May 23, 2006, Fiction, 496 pages. Receiving mysterious clues about a shooting murder in Montreal, Tempe Brennan wonders if the victim may have been a Jewish black market antiquities trader and teams up with .... Deadly Decisions A Novel, Kathy Reichs, Aug 8, 2000, Fiction, 336 pages. Nobody tells a chilling story like international bestselling author Kathy Reichs, whose "most valuable tool is her expertise...she's the real thing" (New York Newsday). Drawing .... Corpi freddi La serie di Temperance Brennan #1, Kathy Reichs, , Fiction, 362 pages. -

Bugs, Bones & Botany Workshop October 30-November 4, 2016 Gainesville, Florida
Bugs, Bones & Botany Workshop October 30‐November 4, 2016 Gainesville, Florida October 30, 2016 Classroom Lecture Topics 1. Entomology: History & Overview of Entomology, Dr. Jason 8AM‐12 PM Byrd 2. Anthropology: Introduction to Forensic Anthropology, Instructor: Dr. Mike Warren 3. Botany: Using Botanical Evidence in a Forensic Investigation, Instructor: Dr. David Hall 4. Crime Scene: Search & Field Recovery Techniques, Gravesite excavation, begin 1‐5 PM Field Topic 1. Entomology: Field Demonstration of Collection Procedures, Instructor: Dr. Jason Byrd 2. Anthropology: Hands‐on Skeletal Analysis Instructor: Dr. Mike Warren 3. Botany: Where Plants Grow & Mapping/Collecting Equipment, Instructor: Dr. David Hall November 1, 2016 Classroom Topic 1. Entomology: Processing a Death Scene for Entomological 8‐12 AM Evidence, Instructor: Dr. Jason Byrd 2. Anthropology: Human and Nonhuman Skeletal Anatomy, Instructor: Dr. Mike Warren 3. Botany: Characteristics of Plants, Instructor: Dr. David Hall 1‐5 PM Field topic 1. Entomology: Collection of Entomological Evidence Instructor: Dr. Jason Byrd 2. Anthropology: Students will excavate buried remains, Instructor: Dr. Mike Warren 3. Botany: Collecting Botanical Evidence & Mapping Surface Scatter, Instructor: Dr. David Hall November 2, 2016 Classroom Topic 1. Entomology: Estimation of PMI Using Entomological Evidence, 8 AM – 12 PM Instructor: Dr. Jason Byrd 2. Anthropology: Methods of Human Identification Instructor: Dr. Mike Warren 3. Botany: When to call a Forensic Botanist Instructor: Dr. David Hall 1‐5 PM Field Topic 1. Entomology: Continue Processing of Entomological Evidence, Instructor: Dr. Jason Byrd 2. Anthropology: Continue Processing Anthropological Evidence, Instructor: Dr. Mike Warren 3. Botany: Continue Processing Botanical Evidence & Surface Scatter, Instructor: Dr. David Hall November 3, 2016 Classroom Topic 1. -

How I Set up My Own Body Farm by Jennifer Dean
How I Set Up My Own Body Farm By Jennifer Dean To prepare for a new forensic science elective at Camas High School, and determined to make this new course an exciting application of biology and chemistry principles, I began by collecting forensic science resources, ordering books and enrolling in the local community college course on forensic science. I spent every extra hour soaking up as much as I could about this field during the “time off” teachers get in the summer. I was particularly fascinated by the fields of forensic entomology and anthropology. From books such as Stiff: The Curious Lives of Human Cadavers by Mary Roach and Dr. Bill Bass’s work around the creation of a human body farm at the University of Tennessee Forensic Anthropology Center, I decided to create something similar for our new high school forensic science program. As we met for dinner after a day of revitalizing workshops in Seattle at an NSTA conference, I shared my thoughts about the creation of an animal body farm with my talented and dedicated colleagues. These teachers have a passion for their students and their work. I felt free to share these ideas with them and know I’d be supported in making it a reality. As the Science Department, we formally agreed to dedicate ourselves to submitting grants to make it a reality. Back at work, I sent copies of the farm proposal to my immediate supervisors, and they responded with letters of support. The next step was getting the farm started—with or without grant money—because the first class of forensic science would be starting in the fall. -

51ST ANNUAL CONVENTION March 5–8, 2020 Boston, MA
Northeast Modern Language Association 51ST ANNUAL CONVENTION March 5–8, 2020 Boston, MA Local Host: Boston University Administrative Sponsor: University at Buffalo SUNY 1 BOARD OF DIRECTORS President Carole Salmon | University of Massachusetts Lowell First Vice President Brandi So | Department of Online Learning, Touro College and University System Second Vice President Bernadette Wegenstein | Johns Hopkins University Past President Simona Wright | The College of New Jersey American and Anglophone Studies Director Benjamin Railton | Fitchburg State University British and Anglophone Studies Director Elaine Savory | The New School Comparative Literature Director Katherine Sugg | Central Connecticut State University Creative Writing, Publishing, and Editing Director Abby Bardi | Prince George’s Community College Cultural Studies and Media Studies Director Maria Matz | University of Massachusetts Lowell French and Francophone Studies Director Olivier Le Blond | University of North Georgia German Studies Director Alexander Pichugin | Rutgers, State University of New Jersey Italian Studies Director Emanuela Pecchioli | University at Buffalo, SUNY Pedagogy and Professionalism Director Maria Plochocki | City University of New York Spanish and Portuguese Studies Director Victoria L. Ketz | La Salle University CAITY Caucus President and Representative Francisco Delgado | Borough of Manhattan Community College, CUNY Diversity Caucus Representative Susmita Roye | Delaware State University Graduate Student Caucus Representative Christian Ylagan | University