And Antagonists Between the Pithed Rabbit and Rat J.M
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
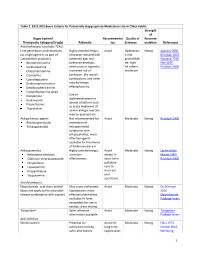
Table 2. 2012 AGS Beers Criteria for Potentially
Table 2. 2012 AGS Beers Criteria for Potentially Inappropriate Medication Use in Older Adults Strength of Organ System/ Recommendat Quality of Recomm Therapeutic Category/Drug(s) Rationale ion Evidence endation References Anticholinergics (excludes TCAs) First-generation antihistamines Highly anticholinergic; Avoid Hydroxyzin Strong Agostini 2001 (as single agent or as part of clearance reduced with e and Boustani 2007 combination products) advanced age, and promethazi Guaiana 2010 Brompheniramine tolerance develops ne: high; Han 2001 Carbinoxamine when used as hypnotic; All others: Rudolph 2008 Chlorpheniramine increased risk of moderate Clemastine confusion, dry mouth, Cyproheptadine constipation, and other Dexbrompheniramine anticholinergic Dexchlorpheniramine effects/toxicity. Diphenhydramine (oral) Doxylamine Use of diphenhydramine in Hydroxyzine special situations such Promethazine as acute treatment of Triprolidine severe allergic reaction may be appropriate. Antiparkinson agents Not recommended for Avoid Moderate Strong Rudolph 2008 Benztropine (oral) prevention of Trihexyphenidyl extrapyramidal symptoms with antipsychotics; more effective agents available for treatment of Parkinson disease. Antispasmodics Highly anticholinergic, Avoid Moderate Strong Lechevallier- Belladonna alkaloids uncertain except in Michel 2005 Clidinium-chlordiazepoxide effectiveness. short-term Rudolph 2008 Dicyclomine palliative Hyoscyamine care to Propantheline decrease Scopolamine oral secretions. Antithrombotics Dipyridamole, oral short-acting* May -
ALPHA ADRENOCEPTORS and HUMAN SEXUAL FUNCTION Alan
8 1995 Elsevier Science B. V. All rights reserved. The Pharmacology of Sexual Function and Dysfunction J. Bancroft, editor 307 ALPHA ADRENOCEPTORS AND HUMAN SEXUAL FUNCTION Alan J Riley Field Place, Dunsmore, Buckinghamshire, HP22 6QH, UK Introduction Sexual functioning involves complex physiological processes which rely on the interplay of many central and peripheral neurotransmitter systems. Disturbances in any one of these systems might be associated with disturbed sexual function which, when recognised, may be alleviated by appropriate pharmacological manipulation, although at the present time this is more hypothesis than reality, The sympathetic nervous system is involved actively at various levels in the normal control of sexual responses. The effects of sympathetic activation are mediated by the release of noradrenaline from nerve terminals and the increased secretion of adrenaline from the adrenal medulla. These catecholamines selectively activate specific cellular sites in target tissues known as adrenoceptors (previously termed adrenergic receptors) to mediate responses. Almost fifty years ago, Alquist realised that tissue responses to catecholamines were mediated through two distinct types of receptors which he designated a and/? [1]. This review focuses on the involvement of a-adrenoceptors in human sexual functioning and dysfunction. Alpha adrenoceptors are located both pre- and post- synaptically and they were classified as either ar or af adrenoceptors according to location; or, being postsynaptic and az presynaptic. This classification continues to be used in some texts. However, as highly specific and selective pharmacological tools became available, this locational subclassification is found not always to be appropriate. Nowadays, classification of a-adrenoceptors is more appropriately based on pharmacological activity and additional subtypes of a-adrenoceptors have been identified by radioligand binding and molecular biological techniques [2]. -

)&F1y3x PHARMACEUTICAL APPENDIX to THE
)&f1y3X PHARMACEUTICAL APPENDIX TO THE HARMONIZED TARIFF SCHEDULE )&f1y3X PHARMACEUTICAL APPENDIX TO THE TARIFF SCHEDULE 3 Table 1. This table enumerates products described by International Non-proprietary Names (INN) which shall be entered free of duty under general note 13 to the tariff schedule. The Chemical Abstracts Service (CAS) registry numbers also set forth in this table are included to assist in the identification of the products concerned. For purposes of the tariff schedule, any references to a product enumerated in this table includes such product by whatever name known. Product CAS No. Product CAS No. ABAMECTIN 65195-55-3 ACTODIGIN 36983-69-4 ABANOQUIL 90402-40-7 ADAFENOXATE 82168-26-1 ABCIXIMAB 143653-53-6 ADAMEXINE 54785-02-3 ABECARNIL 111841-85-1 ADAPALENE 106685-40-9 ABITESARTAN 137882-98-5 ADAPROLOL 101479-70-3 ABLUKAST 96566-25-5 ADATANSERIN 127266-56-2 ABUNIDAZOLE 91017-58-2 ADEFOVIR 106941-25-7 ACADESINE 2627-69-2 ADELMIDROL 1675-66-7 ACAMPROSATE 77337-76-9 ADEMETIONINE 17176-17-9 ACAPRAZINE 55485-20-6 ADENOSINE PHOSPHATE 61-19-8 ACARBOSE 56180-94-0 ADIBENDAN 100510-33-6 ACEBROCHOL 514-50-1 ADICILLIN 525-94-0 ACEBURIC ACID 26976-72-7 ADIMOLOL 78459-19-5 ACEBUTOLOL 37517-30-9 ADINAZOLAM 37115-32-5 ACECAINIDE 32795-44-1 ADIPHENINE 64-95-9 ACECARBROMAL 77-66-7 ADIPIODONE 606-17-7 ACECLIDINE 827-61-2 ADITEREN 56066-19-4 ACECLOFENAC 89796-99-6 ADITOPRIM 56066-63-8 ACEDAPSONE 77-46-3 ADOSOPINE 88124-26-9 ACEDIASULFONE SODIUM 127-60-6 ADOZELESIN 110314-48-2 ACEDOBEN 556-08-1 ADRAFINIL 63547-13-7 ACEFLURANOL 80595-73-9 ADRENALONE -

(12) Patent Application Publication (10) Pub. No.: US 2012/0115729 A1 Qin Et Al
US 201201.15729A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2012/0115729 A1 Qin et al. (43) Pub. Date: May 10, 2012 (54) PROCESS FOR FORMING FILMS, FIBERS, Publication Classification AND BEADS FROM CHITNOUS BOMASS (51) Int. Cl (75) Inventors: Ying Qin, Tuscaloosa, AL (US); AOIN 25/00 (2006.01) Robin D. Rogers, Tuscaloosa, AL A6II 47/36 (2006.01) AL(US); (US) Daniel T. Daly, Tuscaloosa, tish 9.8 (2006.01)C (52) U.S. Cl. ............ 504/358:536/20: 514/777; 426/658 (73) Assignee: THE BOARD OF TRUSTEES OF THE UNIVERSITY OF 57 ABSTRACT ALABAMA, Tuscaloosa, AL (US) (57) Disclosed is a process for forming films, fibers, and beads (21) Appl. No.: 13/375,245 comprising a chitinous mass, for example, chitin, chitosan obtained from one or more biomasses. The disclosed process (22) PCT Filed: Jun. 1, 2010 can be used to prepare films, fibers, and beads comprising only polymers, i.e., chitin, obtained from a suitable biomass, (86). PCT No.: PCT/US 10/36904 or the films, fibers, and beads can comprise a mixture of polymers obtained from a suitable biomass and a naturally S3712). (4) (c)(1), Date: Jan. 26, 2012 occurring and/or synthetic polymer. Disclosed herein are the (2), (4) Date: an. AO. films, fibers, and beads obtained from the disclosed process. O O This Abstract is presented solely to aid in searching the sub Related U.S. Application Data ject matter disclosed herein and is not intended to define, (60)60) Provisional applicationpp No. 61/182,833,sy- - - s filed on Jun. -

Stems for Nonproprietary Drug Names
USAN STEM LIST STEM DEFINITION EXAMPLES -abine (see -arabine, -citabine) -ac anti-inflammatory agents (acetic acid derivatives) bromfenac dexpemedolac -acetam (see -racetam) -adol or analgesics (mixed opiate receptor agonists/ tazadolene -adol- antagonists) spiradolene levonantradol -adox antibacterials (quinoline dioxide derivatives) carbadox -afenone antiarrhythmics (propafenone derivatives) alprafenone diprafenonex -afil PDE5 inhibitors tadalafil -aj- antiarrhythmics (ajmaline derivatives) lorajmine -aldrate antacid aluminum salts magaldrate -algron alpha1 - and alpha2 - adrenoreceptor agonists dabuzalgron -alol combined alpha and beta blockers labetalol medroxalol -amidis antimyloidotics tafamidis -amivir (see -vir) -ampa ionotropic non-NMDA glutamate receptors (AMPA and/or KA receptors) subgroup: -ampanel antagonists becampanel -ampator modulators forampator -anib angiogenesis inhibitors pegaptanib cediranib 1 subgroup: -siranib siRNA bevasiranib -andr- androgens nandrolone -anserin serotonin 5-HT2 receptor antagonists altanserin tropanserin adatanserin -antel anthelmintics (undefined group) carbantel subgroup: -quantel 2-deoxoparaherquamide A derivatives derquantel -antrone antineoplastics; anthraquinone derivatives pixantrone -apsel P-selectin antagonists torapsel -arabine antineoplastics (arabinofuranosyl derivatives) fazarabine fludarabine aril-, -aril, -aril- antiviral (arildone derivatives) pleconaril arildone fosarilate -arit antirheumatics (lobenzarit type) lobenzarit clobuzarit -arol anticoagulants (dicumarol type) dicumarol -

Antihypertensive Agents Using ALZET Osmotic Pumps
ALZET® Bibliography References on the Administration of Antihypertensive Agents Using ALZET Osmotic Pumps 1. Atenolol Q7652: W. B. Zhao, et al. Stimulation of beta-adrenoceptors up-regulates cardiac expression of galectin-3 and BIM through the Hippo signalling pathway. British Journal of Pharmacology 2019;176(14):2465-2481 Agents: Isoproterenol; propranolol; carvedilol; atenolol; ICI-118551 Vehicle: saline; ascorbic acid, buffered; Route: SC; Species: Mice; Pump: 2001; Duration: 1 day; 2 days; 7 days; ALZET Comments: Dose ((ISO 0.6, 6, 20 mg/kg/d), (Prop 2 mg/kg/d), (Carv 2 mg/kg/d), (AT 2 mg/kg/d), (ICI 1 mg/kg/d)); saline with 0.4 mM ascorbic acid used; Controls were non-transgenic and received mp w/ vehicle; animal info (12-16 weeks, Male, (C57BL/6J, beta2-TG, Mst1-TG, or dnMst1-TG)); ICI-118551 is a beta2-antagonist with the structure (2R,3S)-1-[(7-methyl-2,3-dihydro-1H-inden-4-yl)oxy]-3-(propan-2-ylamino)butan-2-ol; cardiovascular; Minipumps were removed to allow for washout of ISO overnight prior to imaging; Q7241: M. N. Nguyen, et al. Mechanisms responsible for increased circulating levels of galectin-3 in cardiomyopathy and heart failure. Sci Rep 2018;8(1):8213 Agents: Isoproterenol, Atenolol, ICI-118551 Vehicle: Saline, ascorbic acid; Route: SC; Species: Mice; Pump: Not Stated; Duration: 48 Hours; ALZET Comments: Dose: ISO (2, 6 or 30 mg/kg/day; atenolol (2 mg/kg/day), ICI-118551 (1 mg/kg/day); 0.4 mM ascorbic used; animal info (12 14 week-old C57Bl/6 mice); cardiovascular; Q6161: C. -

(12) Patent Application Publication (10) Pub. No.: US 2005/0049256A1 Lorton Et Al
US 2005.0049256A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2005/0049256A1 LOrton et al. (43) Pub. Date: Mar. 3, 2005 (54) TREATMENT OF INFLAMMATORY Publication Classification AUTOIMMUNE DISEASES WITH ALPHA-ADRENERGIC ANTAGONSTS AND BETA-ADRENERGICAGONSTS (51) Int. Cl." ...................... A61K 31/517; A61K 31/137 (76) Inventors: Dianne Lorton, Avondale, AZ (US); (52) U.S. Cl. ..................... 514/252.17; 514/649; 514/651 Cheri Lubahn, Glendale, AZ (US) Correspondence Address: JENNINGS, STROUSS & SALMON, P.L.C. (57) ABSTRACT 201 E. WASHINGTON ST, 11TH FLOOR PHOENIX, AZ 85004 (US) The present invention discloses a novel compound and (21) Appl. No.: 10/928,437 method for the treatment of inflammatory autoimmune dis eases, for example, rheumatoid arthritis, using C.-adrenergic (22) Filed: Aug. 27, 2004 antagonists and B-adrenergic agonists in combination. Treat ment of animals, namely humans, with an O-adrenergic Related U.S. Application Data antagonist, preferably, phentolamine, and a B-adrenergic agonist, preferably terbutaline, in combination can Signifi (60) Provisional application No. 60/498.367, filed on Aug. cantly Suppress the joint destruction and inflammation due to 27, 2003. disease in these animals. Patent Application Publication Mar. 3, 2005 Sheet 1 of 5 US 2005/0049256A1 Patent Application Publication Mar. 3, 2005 Sheet 2 of 5 US 2005/0049256A1 e \ s o o (ulu) pA ped)00 IBuedos. IOC Patent Application Publication Mar. 3, 2005 Sheet 3 of 5 US 2005/0049256A1 as o N w N o et N O CY) c5 O t al L cN Y D. a s t is r s re y (uu) supW pedoo Patent Application Publication US 2005/004925.6 A1 |0*>d 0|| eIOOS 3 guide foupe Patent Application Publication Mar. -

Current and Experimental Therapeutics for the Treatment of Opioid Addiction
105 CURRENT AND EXPERIMENTAL THERAPEUTICS FOR THE TREATMENT OF OPIOID ADDICTION PAUL J. FUDALA GEORGE E. WOODY Currently, numerous effective pharmacologic and behav- ing treatment have been addicted to heroin or other opioids ioral therapies are available for the treatment of opioid ad- for 2 to 3 years, some for 30 years or more. Thus, treatment diction, and these two types of therapies often are combined usually involves changes in patients’ lifestyles. Although to optimize patient management. Newer therapeutic op- generally ineffective in producing sustained remission unless tions may take various forms. For example, methadone combined with long-term pharmacologic, psychosocial, or maintenance is an established treatment modality, whereas behavioral therapies, detoxification alone continues to be the use of buprenorphine and naloxone in an office-based widely used and studied. It is sometimes the only option setting represents a new variation on that theme. Clonidine available for patients who do not meet United States Food has been used extensively to ameliorate opioid withdrawal and Drug Administration (FDA) criteria for, do not desire, signs, whereas lofexidine is a structural analogue that ap- or do not have access to agonist medications such as metha- pears to have less hypotensive and sedating effects. The done or methadyl acetate (L-␣-acetylmethadol or LAAM). depot dosage form of naltrexone, currently under develop- The detoxification process may include use of opioid ment, may increase compliance with a medication that has agonists (e.g., methadone), partial agonists (e.g., buprenor- been an effective opioid antagonist but that has been un- phine), antagonists (e.g., naloxone, naltrexone), or nonopi- derused secondary to patient nonacceptance. -

Add-On Effect of Bedtime Dosing of the Α1-Adrenergic Receptor
1097 Hypertens Res Vol.30 (2007) No.11 p.1097-1105 Original Article Add-On Effect of Bedtime Dosing of the α1 -Adrenergic Receptor Antagonist Doxazosin on Morning Hypertension and Left Ventricular Hypertrophy in Patients Undergoing Long-Term Amlodipine Monotherapy Toshio IKEDA1), Tomoko GOMI1), Yuko SHIBUYA1), Shingo SHINOZAKI1), Yoshifumi SUZUKI1), and Nami MATSUDA1) High morning blood pressure is related to target organ damage and future cardiovascular events. Chrono- biologic therapies focusing on the early morning period may be an important strategy for antihypertensive therapy. The aim of this study was to clarify the add-on effects of bedtime dosing of the α1 -adrenergic recep- tor antagonist doxazosin on morning blood pressure in patients with essential hypertension who were under long-acting calcium channel blocker amlodipine monotherapy. The add-on effects of doxazosin at the max- imum dose of 6 mg at bedtime on home blood pressure and left ventricular geometry for 1 year were inves- tigated in 49 subjects (37 men and 12 women, aged 57.5±9.1 years) with morning hypertension who had been treated with amlodipine alone for more than 1 year. Doxazosin induced a significant decrease in morn- ing blood pressure (145.6±5.6/91.5±5.4 to 132.4±3.7/83.6±5.6 mmHg, p≤0.001/<0.001) without a change of evening blood pressure (128.9±5.1/79.8±5.1 to 127.7±6.0/78.8±6.2 mmHg, p=0.056/0.051). Left ventricular mass index (LVMI; 124.8±19.8 to 95.6±15.7 g/m2, p<0.001), relative wall thickness (0.457±0.061 to 0.405±0.047, p<0.001) and homeostasis model assessment of the insulin resistance index (HOMA-IR; 2.62±1.43 to 1.33±0.75, p<0.001) were decreased after doxazosin therapy. -

Initial Medication Selection for Treatment of Hypertension in an Open-Panel HMO
J Am Board Fam Pract: first published as 10.3122/jabfm.8.1.1 on 1 January 1995. Downloaded from Initial Medication Selection For Treatment Of Hypertension In An Open-Panel HMO Micky jerome, PharmD, MBA, George C. Xakellis, MD, Greg Angstman, MD, and Wayne Patchin, MBA Background: During the past 25 years recommendations for treating hypertension have evolved from a stepped-care approach to monotherapy or sequential monotherapy as experience has been gained and new antihypertensive agents have been introduced. In an effort to develop a disease management strategy for hypertension, we investigated the prescribing patterns of initial medication therapy for newly treated hypertensive patients. Methods: We examined paid claims data of an open-panel HMO located in the midwest. Charts from 377 patients with newly treated hypertension from a group of 12,242 hypertenSive patients in a health insurance population of 85,066 persons were studied. The type of medication regimen received by patients newly treated for hypertension during an 18-month period was categorized into monotherapy, sequential monotherapy, stepped care, and initial treatment with multiple agents. With monotherapy, the class of medication was also reported. Associations between use of angiotensin-converting enzyme (ACE) inhibitors, calcium channel blockers, or (3-blockers and presence of comorbid conditions were reported. Results: Fifty-five percent of patients received monotherapy, 22 percent received stepped care, and 18 percent received sequential monotherapy. Of those 208 patients receiving monotherapy, 30 percent were prescribed a calcium channel blocker, 22 percent an ACE inhibitor, and 14 percent a f3-blocker. No customization of treatment for comorbid conditions was noted. -

Long Acting, Reversible Veterinary Sedative and Analgesic And
University of Kentucky UKnowledge Veterinary Science Faculty Patents Veterinary Science 7-11-2006 Long Acting, Reversible Veterinary Sedative and Analgesic and Method of Use Thomas Tobin University of Kentucky, [email protected] Right click to open a feedback form in a new tab to let us know how this document benefits oy u. Follow this and additional works at: https://uknowledge.uky.edu/gluck_patents Part of the Veterinary Medicine Commons Recommended Citation Tobin, Thomas, "Long Acting, Reversible Veterinary Sedative and Analgesic and Method of Use" (2006). Veterinary Science Faculty Patents. 14. https://uknowledge.uky.edu/gluck_patents/14 This Patent is brought to you for free and open access by the Veterinary Science at UKnowledge. It has been accepted for inclusion in Veterinary Science Faculty Patents by an authorized administrator of UKnowledge. For more information, please contact [email protected]. US007074834B2 (12) United States Patent (10) Patent N0.: US 7,074,834 B2 Tobin (45) Date of Patent: Jul. 11, 2006 (54) LONG ACTING, REVERSIBLE VETERINARY 4,742,054 A 5/1988 Naftchi SEDATIVE AND ANALGESIC AND METHOD 4,950,648 A 8/1990 Raddatz et a1. OF USE 5,635,204 A * 6/1997 GevirtZ et a1. ............ .. 424/449 5,942,241 A 8/1999 Chasin et a1. (75) Inventor: Thomas Tobin, Lexington, KY (US) 5,958,933 A 9/1999 Naftchi (73) Assignee: University of Kentucky Foundation, OTHER PUBLICATIONS Lexington, KY (US) MEDLINE AN 20000025586, Veveris et al, Brit. J. Pharmacol, 128 (5), 1089-97, Nov. 1999, abstract.* ( * ) Notice: Subject to any disclaimer, the term of this Veterinary Pharmacology and Therapeutics, Adams, pp. -

And Α2-Adrenoceptor Subtypes in the Vasopressor Responses Induced by Dihydroergotamine in Ritanserin-Pretreated Pithed Rats Eduardo Rivera-Mancilla1, Victor H
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Erasmus University Digital Repository Rivera-Mancilla et al. The Journal of Headache and Pain (2017) 18:104 The Journal of Headache DOI 10.1186/s10194-017-0812-4 and Pain RESEARCHARTICLE Open Access The role of α1- and α2-adrenoceptor subtypes in the vasopressor responses induced by dihydroergotamine in ritanserin-pretreated pithed rats Eduardo Rivera-Mancilla1, Victor H. Avilés-Rosas1, Guadalupe Manrique-Maldonado1, Alain H. Altamirano-Espinoza1, Belinda Villanueva-Castillo1, Antoinette MaassenVanDenBrink2 and Carlos M. Villalón1* Abstract Background: Dihydroergotamine (DHE) is an acute antimigraine agent that displays affinity for dopamine D2-like receptors, serotonin 5-HT1/2 receptors and α1/α2-adrenoceptors. Since activation of vascular α1/α2-adrenoceptors results in systemic vasopressor responses, the purpose of this study was to investigate the specific role of α1- and α2-adrenoceptors mediating DHE-induced vasopressor responses using several antagonists for these receptors. Methods: For this purpose, 135 male Wistar rats were pithed and divided into 35 control and 100 pretreated i.v. with ritanserin (100 μg/kg; to exclude the 5-HT2 receptor-mediated systemic vasoconstriction). Then, the vasopressor responses to i.v. DHE (1–3100 μg/kg, given cumulatively) were determined after i.v. administration of some α1/α2-adrenoceptor antagonists. Results: In control animals (without ritanserin pretreatment), the vasopressor responses to DHE were: (i) unaffected after prazosin (α1;30μg/kg); (ii) slightly, but significantly, blocked after rauwolscine (α2;300μg/kg); and (iii) markedly blocked after prazosin (30 μg/kg) plus rauwolscine (300 μg/kg).