Defective Color Vision and Its Inheritance by George Wald*
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Unrestricted Immigration and the Foreign Dominance Of
Unrestricted Immigration and the Foreign Dominance of United States Nobel Prize Winners in Science: Irrefutable Data and Exemplary Family Narratives—Backup Data and Information Andrew A. Beveridge, Queens and Graduate Center CUNY and Social Explorer, Inc. Lynn Caporale, Strategic Scientific Advisor and Author The following slides were presented at the recent meeting of the American Association for the Advancement of Science. This project and paper is an outgrowth of that session, and will combine qualitative data on Nobel Prize Winners family histories along with analyses of the pattern of Nobel Winners. The first set of slides show some of the patterns so far found, and will be augmented for the formal paper. The second set of slides shows some examples of the Nobel families. The authors a developing a systematic data base of Nobel Winners (mainly US), their careers and their family histories. This turned out to be much more challenging than expected, since many winners do not emphasize their family origins in their own biographies or autobiographies or other commentary. Dr. Caporale has reached out to some laureates or their families to elicit that information. We plan to systematically compare the laureates to the population in the US at large, including immigrants and non‐immigrants at various periods. Outline of Presentation • A preliminary examination of the 609 Nobel Prize Winners, 291 of whom were at an American Institution when they received the Nobel in physics, chemistry or physiology and medicine • Will look at patterns of -

Sensitivities in Older Eyes with Good Acuity: Cross-Sectional Norms
Sensitivities in Older Eyes With Good Acuity: Cross-Sectional Norms Alvin Eisner,*f Susan A. Fleming,*! Michael L. Kleins and W. Manning Mauldinf We measured several indices of foveal visual function for a large group of people aged 60 and older. The data reported in this paper are from individuals who had good acuity in each eye and met a number of other criteria for good ocular health. For each index, we described the rate of cross-sectional change with age using linear regression statistics. We found age-related change for eyes having 20/20 or better acuity to exist for several different indices. Sensitivity mediated by the blue-sensitive cones decreased with age, as expected. However, the rate of decrease was faster for females than for males. At least part of the difference was associated with different rates of lenticular change. Absolute sensitivity at long wavelengths also decreased with age, but at the same rate for each sex. Rayleigh color matches changed with age in a manner consistent with underlying age-related decreases of effective foveal cone photopigment density. However, not all indices showed age-dependent changes. For instance, the time constant describing the rate of photopic dark adaptation did not appear to change with age. Invest Ophthalmol Vis Sci 28:1824-1831, 1987 Human visual function changes with age. Cross- criteria for good ocular health in each eye. In particu- sectional age-related changes have been reported for lar, we have tested people having good acuity in each Snellen acuity,1 spatial2"4 and temporal4 contrast sen- eye. -

ERG), Molecular and Behavioral Studies
Vision Research 38 (1998) 3377–3385 Severity of color vision defects: electroretinographic (ERG), molecular and behavioral studies M.A. Crognale a,b,*, D.Y. Teller a,c, A.G. Motulsky d,e, S.S. Deeb d,e a Department of Psychology, Uni6ersity of Washington, Box 351525, Seattle, WA 98195-1525, USA b Department of Ophthalmology, Uni6ersity of Washington, Seattle, WA, USA c Department of Physiology/Biophysics, Uni6ersity of Washington, Seattle, WA, USA d Department of Genetics, Uni6ersity of Washington, Seattle, WA, USA e Department of Medicine, Uni6ersity of Washington, Seattle, WA, USA Received 10 July 1997; received in revised form 29 October 1997 Abstract Earlier research on phenotype/genotype relationships in color vision has shown imperfect predictability of color matching from the photopigment spectral sensitivities inferred from molecular genetic analysis. We previously observed that not all of the genes of the X-chromosome linked photopigment gene locus are expressed in the retina. Since sequence analysis of DNA does not necessarily reveal which of the genes are expressed into photopigments, we used ERG spectral sensitivities and adaptation measurements to assess expressed photopigment complement. Many deuteranomalous subjects had L, M, and L–M hybrid genes. The ERG results showed that M pigment is not present in measurable quantities in deutan subjects. Using these results to determine gene expression improved the correlations between inferred pigment separation and color matching. Furthermore, we found a subject who had normal L and M genes and normal proximal promoter sequences, yet he had a single photopigment (M) by ERG and tested as a protanope. These results demonstrate the utility of ERG measurements in studies of molecular genetics of color vision deficiencies, and further support the conclusion that not all genes are expressed in color deficient subjects. -

Color Blindness LEVELED BOOK • W a Reading A–Z Level W Leveled Book Word Count: 1,349 Color Blindness Connections Writing Choose Two Forms of Color Blindness
Color Blindness LEVELED BOOK • W A Reading A–Z Level W Leveled Book Word Count: 1,349 Color Blindness Connections Writing Choose two forms of color blindness. Write a report that compares the two conditions and their effects on a person’s life. Math Research the statistics about the number of people with the various forms of color blindness in your country. Organize your results in a pie chart. • W Q •T Written by Cheryl Reifsnyder Visit www.readinga-z.com for thousands of books and materials. www.readinga-z.com Words to Know Color ancestry molecules complementary photopigments cone cells prism Blindness defects retina genetic disorder rod cells hereditary wavelengths Photo Credits: Front cover, Back cover: © pretoperola/123RF; title page (Both): © shironosov/ iStock/Thinkstock; page 3: © Véronique Burger/Science Source; page 4 (Both): © Michael Shake/Dreamstime.com; page 5: © Designua/Dreamstime.com; page 6 (top): © Gunilla Elam/Science Source; page 6 (Bottom): © Steve Gschmeissner/Science Source; page 7 (Both): © Ted Kinsman/ Science Source; page 8 (left, Both): © anatchant/iStock/Thinkstock; page 8 (center, Both): © Nadezhda Bolotina/Hemera/Thinkstock; page 8 (right, Both): © iremphotography/iStock/Thinkstock; page 9 (all): © Popartic/iStock Editorial/Thinkstock; page 10: © RVN/Alamy Stock Photo; page 12: © Alexander Kaludov/123RF; page 13: © EnChroma; page 14: © Neitz LaBoratory; page 15: © Phanie/Alamy Stock Photo Written by Cheryl Reifsnyder www.readinga-z.com Focus Question Color Blindness Level W Leveled Book Correlation © Learning A–Z LEVEL W Written By Cheryl Reifsnyder What causes color blindness, and how Fountas & Pinnell S can it affect a person’s life? All rights reserved. -
RETINAL DISORDERS Eye63 (1)
RETINAL DISORDERS Eye63 (1) Retinal Disorders Last updated: May 9, 2019 CENTRAL RETINAL ARTERY OCCLUSION (CRAO) ............................................................................... 1 Pathophysiology & Ophthalmoscopy ............................................................................................... 1 Etiology ............................................................................................................................................ 2 Clinical Features ............................................................................................................................... 2 Diagnosis .......................................................................................................................................... 2 Treatment ......................................................................................................................................... 2 BRANCH RETINAL ARTERY OCCLUSION ................................................................................................ 3 CENTRAL RETINAL VEIN OCCLUSION (CRVO) ..................................................................................... 3 Pathophysiology & Etiology ............................................................................................................ 3 Clinical Features ............................................................................................................................... 3 Diagnosis ......................................................................................................................................... -

The Genetics of Normal and Defective Color Vision
Vision Research xxx (2011) xxx–xxx Contents lists available at ScienceDirect Vision Research journal homepage: www.elsevier.com/locate/visres Review The genetics of normal and defective color vision Jay Neitz ⇑, Maureen Neitz University of Washington, Dept. of Ophthalmology, Seattle, WA 98195, United States article info a b s t r a c t Article history: The contributions of genetics research to the science of normal and defective color vision over the previ- Received 3 July 2010 ous few decades are reviewed emphasizing the developments in the 25 years since the last anniversary Received in revised form 25 November 2010 issue of Vision Research. Understanding of the biology underlying color vision has been vaulted forward Available online xxxx through the application of the tools of molecular genetics. For all their complexity, the biological pro- cesses responsible for color vision are more accessible than for many other neural systems. This is partly Keywords: because of the wealth of genetic variations that affect color perception, both within and across species, Color vision and because components of the color vision system lend themselves to genetic manipulation. Mutations Cone photoreceptor and rearrangements in the genes encoding the long, middle, and short wavelength sensitive cone pig- Colorblindness Cone mosaic ments are responsible for color vision deficiencies and mutations have been identified that affect the Opsin genes number of cone types, the absorption spectra of the pigments, the functionality and viability of the cones, Evolution and the topography of the cone mosaic. The addition of an opsin gene, as occurred in the evolution of pri- Comparative color vision mate color vision, and has been done in experimental animals can produce expanded color vision capac- Cone photopigments ities and this has provided insight into the underlying neural circuitry. -
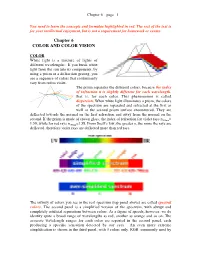
Chapter 6 COLOR and COLOR VISION
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

Genes in Eyecare Geneseyedoc 3 W.M
Genes in Eyecare geneseyedoc 3 W.M. Lyle and T.D. Williams 15 Mar 04 This information has been gathered from several sources; however, the principal source is V. A. McKusick’s Mendelian Inheritance in Man on CD-ROM. Baltimore, Johns Hopkins University Press, 1998. Other sources include McKusick’s, Mendelian Inheritance in Man. Catalogs of Human Genes and Genetic Disorders. Baltimore. Johns Hopkins University Press 1998 (12th edition). http://www.ncbi.nlm.nih.gov/Omim See also S.P.Daiger, L.S. Sullivan, and B.J.F. Rossiter Ret Net http://www.sph.uth.tmc.edu/Retnet disease.htm/. Also E.I. Traboulsi’s, Genetic Diseases of the Eye, New York, Oxford University Press, 1998. And Genetics in Primary Eyecare and Clinical Medicine by M.R. Seashore and R.S.Wappner, Appleton and Lange 1996. M. Ridley’s book Genome published in 2000 by Perennial provides additional information. Ridley estimates that we have 60,000 to 80,000 genes. See also R.M. Henig’s book The Monk in the Garden: The Lost and Found Genius of Gregor Mendel, published by Houghton Mifflin in 2001 which tells about the Father of Genetics. The 3rd edition of F. H. Roy’s book Ocular Syndromes and Systemic Diseases published by Lippincott Williams & Wilkins in 2002 facilitates differential diagnosis. Additional information is provided in D. Pavan-Langston’s Manual of Ocular Diagnosis and Therapy (5th edition) published by Lippincott Williams & Wilkins in 2002. M.A. Foote wrote Basic Human Genetics for Medical Writers in the AMWA Journal 2002;17:7-17. A compilation such as this might suggest that one gene = one disease. -

OCULAR GENE THERAPY TRIALS ADVANCE Results of the First Phase 3 Trial Were Announced
OCULAR GENE THERAPY TRIALS ADVANCE Results of the first phase 3 trial were announced. BY ARON SHAPIRO INNOVATIONS IN RETINA INNOVATIONS “We used to think that our fate was in our AAV FACILITATES GENE DELIVERY stars, but now we know that, in large measure, The nonpathogenic adeno-associated virus (AAV) has our fate is in our genes.” to date been a safe and effective vector for gene delivery. –James Watson1 The recombinant AAV (rAAV) vector has demonstrated increased specificity and efficiency in ocular AAV-mediated Genetic alterations are known to be respon- gene therapy interventions. Recombinant AAV2 (rAAV2) sible for numerous diseases, and so it follows vectors used for gene therapy are derived from the wild-type logically that the best cures for these diseases virus by deleting the entire viral coding region and replac- might lie in correcting genetic anomalies by the process of ing it with the reporter or therapeutic transgene. Combined gene therapy. This approach involves the introduction of AAV serotypes have also been developed. This diversity genes into existing cells in attempts to prevent or cure a of serotypes may lead to more specific and more efficient wide range of diseases previously thought to be incurable.1 transduction. However, successful and efficient transfection Posterior segment disorders are challenging to treat, and of particular cell types in the eye still depends on other fac- current therapies have numerous shortcomings. Many are tors, such as the AAV titer, the site of injection, the amount invasive, run the risk of complications, offer only short-term of passenger DNA, and the specific gene promoters where relief from symptoms, or are unable to directly treat vision transcription initiation takes place.2 loss. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

1 Human Color Vision
CAMC01 9/30/04 3:13 PM Page 1 1 Human Color Vision Color appearance models aim to extend basic colorimetry to the level of speci- fying the perceived color of stimuli in a wide variety of viewing conditions. To fully appreciate the formulation, implementation, and application of color appearance models, several fundamental topics in color science must first be understood. These are the topics of the first few chapters of this book. Since color appearance represents several of the dimensions of our visual experience, any system designed to predict correlates to these experiences must be based, to some degree, on the form and function of the human visual system. All of the color appearance models described in this book are derived with human visual function in mind. It becomes much simpler to understand the formulations of the various models if the basic anatomy, physiology, and performance of the visual system is understood. Thus, this book begins with a treatment of the human visual system. As necessitated by the limited scope available in a single chapter, this treatment of the visual system is an overview of the topics most important for an appreciation of color appearance modeling. The field of vision science is immense and fascinating. Readers are encouraged to explore the liter- ature and the many useful texts on human vision in order to gain further insight and details. Of particular note are the review paper on the mechan- isms of color vision by Lennie and D’Zmura (1988), the text on human color vision by Kaiser and Boynton (1996), the more general text on the founda- tions of vision by Wandell (1995), the comprehensive treatment by Palmer (1999), and edited collections on color vision by Backhaus et al. -

Color Vision Deficiency
Color Vision Deficiency What is color vision deficiency? Color vision deficiency is called “color blindness” by mistake. Actually, the term describes a number of different problems people have with color vision. Abnormal color vision may vary from not being able to tell certain colors apart to not being able to identify any color. Whom does color vision deficiency affect? An estimated 8% of males and fewer than 1% of females have color vision problems. Most color vision problems run in families and are inherited and present at birth. A child inherits a color vision deficiency by receiving a faulty color vision gene from a parent. Abnormal color vision is found in a recessive gene on the X chromosome. Men are born with just one X and one Y chromosome. However, women have two X chromosomes. Because of this, women can sometimes overcome the faulty gene with their second normal X chromosome. Men, unfortunately, do not have a second X chromosome to help compensate for the faulty color vision gene. Heredity does not cause all color vision problems. One common problem happens from the normal aging of the eye’s lens. The lens is clear at birth, but the aging process causes it to darken and yellow. Older adults may have problems identifying certain dark colors, particularly blues. Certain medications as well as inherited or acquired retinal and optic nerve disease, may also affect normal color vision. Who should be tested for color deficiency? Any child who is having difficulty in school should be checked for possible visual problems including color vision impairment.