The Influence of Training Procedures on Generalization Performance in Scent-Detection Rats
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Empty Discarded Pack Data and the Prevalence of Illicit Trade in Cigarettes in California
Pepperdine University Pepperdine Digital Commons School of Public Policy Working Papers School of Public Policy 1-22-2019 Empty Discarded Pack Data and the Prevalence of Illicit Trade in Cigarettes in California James Prieger Pepperdine University, [email protected] Follow this and additional works at: https://digitalcommons.pepperdine.edu/sppworkingpapers Part of the Econometrics Commons, Health Economics Commons, Political Economy Commons, and the Public Affairs, Public Policy and Public Administration Commons Recommended Citation Prieger, James, "Empty Discarded Pack Data and the Prevalence of Illicit Trade in Cigarettes in California" (2019). Pepperdine University, School of Public Policy Working Papers. Paper 75. https://digitalcommons.pepperdine.edu/sppworkingpapers/75 This Article is brought to you for free and open access by the School of Public Policy at Pepperdine Digital Commons. It has been accepted for inclusion in School of Public Policy Working Papers by an authorized administrator of Pepperdine Digital Commons. For more information, please contact [email protected], [email protected], [email protected]. PEPPERDINE UNIVERSITY Empty Discarded Pack Data and the Prevalence of Illicit Trade in Cigarettes in California January 22, 2019 Jonathan Kulick James E. Prieger JDK Analysis Professor 30 Regent #318 Pepperdine University Jersey City, NJ 07302 School of Public Policy 24255 Pacific Coast Highway [email protected] Malibu, CA 90263-7490 James.Prieger@ pepperdine.edu School of Public Policy 24255 Pacific Coast Highway Malibu, CA 90263- 7490 Abstract Illicit trade in tobacco products (ITTP) creates many harms including reduced tax revenues; damages to the economic interests of legitimate actors; funding for organized-crime and terrorist groups; negative effects of participation in illicit markets, such as violence and incarceration; and reduced effectiveness of smoking-reduction policies, leading to increased damage to health. -

Annual Report 2019 CONTENT
中煙國際 (香港) 有限公司 China Tobacco International (HK) Company Limited (Incorporated in Hong Kong with limited liability) Stock code: 6055 Annual Report 2019 CONTENT Definitions 2 Company Information 4 Financial Highlights 5 Chairman’s Statement 6 Management Discussion and Analysis 7 Review of Continuing Connected Transactions 16 Directors and Senior Management 30 Report of the Directors 37 Corporate Governance Report 48 Environmental, Social and Governance Report 2019 60 Independent Auditor’s Report 86 Statement of Profit or Loss and Other Comprehensive Income 91 Statement of Financial Position 92 Statement of Changes in Equity 94 Statement of Cash Flows 95 Notes to the Financial Statements 96 Financial Summary 140 2 China Tobacco International (HK) Company Limited Definitions In this annual report, unless the context otherwise requires, the following terms shall have the meanings set forth below. “AGM” annual general meeting of the Company; “Audit Committee” the audit committee of the Board; “Board” or “Board of Directors” the board of Directors of the Company; “China” or “PRC” the People’s Republic of China; for the purpose of this report only, references to “China” or the “PRC” do not include the Hong Kong Special Administrative Region, the Macau Special Administrative Region and Taiwan; “CNTC” China National Tobacco Corporation* (中國煙草總公司), an enterprise incorporated in the PRC and the ultimate controlling shareholder of the Company; “CNTC Group” CNTC and its subsidiaries; “Company” or “we” China Tobacco International (HK) Company Limited -
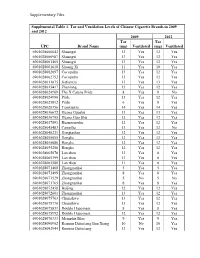
Supplementary Files Supplemental Table 1: Tar And
Supplementary Files Supplemental Table 1: Tar and Ventilation Levels of Chinese Cigarette Brands in 2009 and 2012 2009 2012 Tar Tar UPC Brand Name (mg) Ventilated (mg) Ventilated 6901028000642 Shuangxi 13 Yes 12 Yes 6901028000987 Shuangxi 13 Yes 12 Yes 6901028001465 Shuangxi 13 Yes 12 Yes 6901028001618 Shuang Xi 11 Yes 10 Yes 6901028002097 Cocopalm 13 Yes 12 Yes 6901028002752 Cocopalm 13 Yes 12 Yes 6901028011075 Jiatianxia 13 Yes 13 Yes 6901028015417 Zhenlong 13 Yes 12 Yes 6901028024969 The X Catena Pride 8 Yes 8 No 6901028024990 Pride 13 Yes 12 Yes 6901028025812 Pride 6 Yes 8 Yes 6901028028356 Tianxiaxiu 14 Yes 14 Yes 6901028036672 Huang Guoshu 13 Yes 13 Yes 6901028036795 Huang Guo Shu 13 Yes 12 Yes 6901028037891 Huangguoshu 12 Yes 12 Yes 6901028045483 Camellia 13 Yes 12 No 6901028048231 Hongtashan 12 Yes 12 Yes 6901028055055 Honghe 12 Yes 13 Yes 6901028055086 Honghe 12 Yes 12 Yes 6901028055208 Honghe 12 Yes 12 Yes 6901028065078 Lan zhou 13 Yes 8 Yes 6901028065399 Lan zhou 13 Yes 8 Yes 6901028065580 Lan zhou 11 Yes 8 Yes 6901028071468 Zhongnanhai 5 Yes 5 Yes 6901028071499 Zhongnanhai 8 Yes 8 Yes 6901028071529 Zhongnanhai 5 No 5 No 6901028071765 Zhongnanhai 8 Yes 8 Yes 6901028072458 Beijing 12 Yes 12 Yes 6901028072601 Zhongnanhai 13 Yes 12 Yes 6901028075763 Chunghwa 13 Yes 12 Yes 6901028075770 Chunghwa 13 Yes 12 Yes 6901028075831 Double Happiness 8 Yes 8 Yes 6901028075992 Double Happiness 12 Yes 12 Yes 6901028076333 Memphis Blue 9 Yes 9 Yes 6901028090902 Renmin Dahuitang Ben Xiang 10 No 10 Yes 6901028092944 Renmin Dahuitang 13 Yes 12 Yes -

Fire Standard Compliant Cigarette Brands & Styles
THE STATE OF UTAH Fire Standard Compliant Cigarette Brands and Styles Master Approval List List Design: First Section - Deletions to the list in the last 30 Days Second Section - Additions/Re-Certifications in the last 30 Days Third Section - Main List - Reflects Current Status Updated October 15, 2018 FIRE STANDARD COMPLIANT CIGARETTE BRANDS & STYLES MASTER APPROVAL LIST Page 1 BRANDS NOT RENEWED OR DELETED MANUFACTURER FLAVOR PACKAGE EXPIRATION DATE THIS REPORT NONE BRANDS ADDED OR RECERTIFIED THIS MANUFACTURER FLAVOR PACKAGE CERTIFICATION DATE REPORT Montego Red King Box Liggett Group Inc. Non-Menthol Filter Hard Pack 9/6/2019 Montegi Red 100's Box Liggett Group Inc. Non-Menthol Filter Hard Pack 9/5/2019 Montego Blue 100's Box Liggett Group Inc. Non-Menthol Filter Hard Pack 9/12/2019 Montego Blue Kings Box Liggett Group Inc. Non-Menthol Filter Hard Pack 9/11/2019 Montego Orange 100's Box Liggett Group Inc. Non-Menthol Filter Hard Pack 9/10/2019 Montego Menthol Gold King Liggett Group Inc. Menthol Filter Hard Pack 9/6/2019 Montego Menthol Gold 100's Liggett Group Inc. Menthol Filter Hard Pack 9/5/2019 Montego Menthol Silver King Liggett Group Inc. Menthol Filter Hard Pack 9/11/2019 Montego Menthol Silver 100's Liggett Group Inc. Menthol Filter Hard Pack 9/12/2019 Teton #18 Blue Kings Wind River Tobacco., LLC NS Filter Hard Pack 8/25/2017 Teton #18 Green Menthol Kings Wind River Tobacco., LLC Menthol Filter Hard Pack 11/30/2017 Teton #18 Yellow Kings Wind River Tobacco., LLC NS Filter Hard Pack 9/1/2017 Teton #18 Red 100's Wind River Tobacco., LLC NS Filter Hard Pack 11/27/2017 Teton #6 Gold 100's Wind River Tobacco., LLC NS Filter Hard Pack 11/28/2017 Teton #6 Green Menthol 100's Wind River Tobacco., LLC Menthol Filter Hard Pack 11/29/2017 Teton #6 Black Menthol 100's Wind River Tobacco., LLC Menthol Filter Hard Pack 11/29/2017 Vantage Classic Menthol 100's Soft Pack R.J. -

Utah Cigarette and RYO Authorized Brands, Pub 51
Utah Approved Tobacco Manufacturers and Cigarette/RYO Brands Utah State Tax Commission Publication 51 Effective 08/01/2017 This publication lists all tobacco products certified by their manufacturers as produced for sale in the State of Utah. Products not appearing on this list are prohibited, may not be stamped, and are subject to confiscation as contraband. Even though this publication lists a product, the State of Utah may: · Seek payment under the Tobacco Master Settlement Agreement, · Enforce deposits into a qualified nonparticipating manufacturer escrow account, · Remove a company name or brand family from this publication, or · Take any other legal action to enforce cigarette and tobacco laws. Fire-safe Certification A listed brand includes all styles of that brand unless otherwise indicated. Brands which are followed by an asterisk * are fire-safe certified. For information on whether a specific style of a brand family has been certified fire-safe, please refer to the fire marshal's listing of fire-safe certified cigarettes. This information can be found at http://firemarshal.utah.gov/cigarette-fire-safety-program/ or by clicking on Fire Standard Compliant Cigarette Master List below. Fire Standard Compliant Cigarette Master List Highlighted brands are manufactured by non-participating manufacturers which require special reporting and the NPM (purple) stamp to be affixed. New listed Cigarettes The next scheduled update of this publication is September 1, 2017. Brand Manufacturer New listed RYO The next scheduled update of this publication is September 1, 2017. Brand Manufacturer Delisted Cigarettes The next scheduled update of this publication is September 1, 2017. Brand Manufacturer Pub 51 Page 1 of 4 08/01/2017 Delisted RYO The next scheduled update of this publication is September 1, 2017. -

Illicit Tobacco in New Zealand, 2018 Full Year Report
Illicit tobacco in New Zealand 2018 Full Year Report 24 May 2019 ______ kpmg.com/uk KPMG LLP Tel +44 (0)20 7311 1000 Strategy Group Fax +44 (0)20 7311 3311 15 Canada Square DX 157460 Canary Wharf 5 Canary Wharf London E14 5GL United Kingdom 24 May 2019 Important notice This presentation of key findings (the ‘Report’) has been prepared by KPMG LLP in the UK (‘KPMG UK’) for Philip Morris (New Zealand) Limited and Imperial Tobacco New Zealand Limited, described together in this Important Notice and in this Report as the ‘Beneficiary’, on the basis set out in a private contract dated 18th December 2018 agreed separately with the Beneficiaries. Nothing in this Report constitutes legal advice. Information sources, the scope of our work, and scope and source limitations, are set out in the Appendices to this Report. The scope of our review of the contraband, counterfeit and unbranded segments of the tobacco market within New Zealand was fixed by agreement with the Beneficiaries and is set out in the Appendices. We have satisfied ourselves, so far as possible, that the information presented in this Report is consistent with our information sources but we have not sought to establish the reliability of the information sources by reference to other evidence. This Report has not been designed to benefit anyone except the Beneficiaries. In preparing this Report we have not taken into account the interests, needs or circumstances of anyone apart from the Beneficiaries, even though we have been aware that others might read this Report. This Report is not suitable to be relied on by any party wishing to acquire rights or assert any claims against KPMG LLP (other than the Beneficiaries) for any purpose or in any context. -

Illicit Tobacco in Australia
Illicit tobacco in Australia 2016 Full Year Report 20 March 2017 © 2017 KPMG LLP, a UK limited liability partnership and a member firm of the KPMG network of independent member firms 0 affiliated with KPMG International Cooperative (“KPMG International”), a Swiss entity. All rights reserved. KPMG LLP Tel +44 (0) 20 7311 1000 Strategy Group Fax +44 (0) 20 7311 3311 15 Canada Square DX 157460 Canary Wharf 5 Canary Wharf London E14 5GL United Kingdom 20 March 2017 Important notice This presentation of key findings (the ‘Report’) has been prepared by KPMG LLP in the UK (‘KPMG UK’) for Philip Morris Limited and Imperial Tobacco Australia Limited, described together in this Important Notice and in this Report as the ‘Beneficiaries’, on the basis set out in a private contract dated 16 September 2016 agreed separately with the Beneficiaries. Nothing in this Report constitutes legal advice. Information sources, the scope of our work, and scope and source limitations, are set out in the Appendices to this Report. The scope of our review of the contraband, counterfeit and unbranded segments of the tobacco market within Australia was fixed by agreement with the Beneficiaries and is set out in the Appendices. We have satisfied ourselves, so far as possible, that the information presented in this Report is consistent with our information sources but we have not sought to establish the reliability of the information sources by reference to other evidence. This Report has not been designed to benefit anyone except the Beneficiaries. In preparing this Report we have not taken into account the interests, needs or circumstances of anyone apart from the Beneficiaries, even though we have been aware that others might read this Report. -

2019 Full Year Report
Illicit tobacco in New Zealand 2019 Full Year Report 26 May 2020 kpmg.com/uk KPMG LLP Tel +44 (0)20 7311 1000 Strategy Group Fax +44 (0)20 7311 3311 15 Canada Square DX 157460 Canary Wharf 5 Canary Wharf London E14 5GL United Kingdom 26 May 2020 Important notice This presentation of key findings (the ‘Report’) has been prepared by KPMG LLP in the UK (‘KPMG UK’) for Imperial Tobacco New Zealand Limited, described in this Important Notice and in this Report as the ‘Beneficiary’, on the basis set out in a private contract dated 1st August 2019 agreed separately with the Beneficiary. Nothing in this Report constitutes legal advice. Information sources, the scope of our work, and scope and source limitations, are set out in the Appendices to this Report. The scope of our review of the contraband, counterfeit and unbranded segments of the tobacco market within New Zealand was fixed by agreement with the Beneficiary and is set out in the Appendices. We have satisfied ourselves, so far as possible, that the information presented in this Report is consistent with our information sources but we have not sought to establish the reliability of the information sources by reference to other evidence. This Report has not been designed to benefit anyone except the Beneficiary. In preparing this Report we have not taken into account the interests, needs or circumstances of anyone apart from the Beneficiary, even though we have been aware that others might read this Report. This Report is not suitable to be relied on by any party wishing to acquire rights or assert any claims against KPMG LLP (other than the Beneficiary) for any purpose or in any context. -

Early Evidence About the Predicted Unintended
Open Access Research Early evidence about the predicted unintended consequences of standardised packaging of tobacco products in Australia: a cross-sectional study of the place of purchase, regular brands and use of illicit tobacco Michelle Scollo, Meghan Zacher, Sarah Durkin, Melanie Wakefield To cite: Scollo M, Zacher M, ABSTRACT Strengths and limitations of this study Durkin S, et al. Early Objectives: To test for early evidence whether, following evidence about the predicted the standardisation of tobacco packaging, smokers in ▪ unintended consequences of The dual-frame (landline and mobile phone) Australia were—as predicted by the tobacco industry— standardised packaging of sample design used in this study obtains more rep- tobacco products in Australia: less likely to purchase from small mixed business resentative samples than online or landline-only a cross-sectional study of the retailers, more likely to purchase cheap brands imported phone surveys. place of purchase, regular from Asia and more likely to use illicit tobacco. ▪ A limitation is that recent purchase of potentially brands and use of illicit Design: Serial cross-sectional population telephone illicit branded (contraband) tobacco was only tobacco. BMJ Open 2014;4: surveys in November 2011 (a year prior to assessed in one survey year (2013), and there- e005873. doi:10.1136/ implementation), 2012 (during roll-out) and 2013 (a year fore we could not determine whether it had bmjopen-2014-005873 after implementation). increased between 2011 and 2013. Setting/participants: Smokers aged 18 years and over ▪ As with other surveys, respondent error and mis- ▸ Prepublication history for identified in an annual population survey in the Australian reporting may have affected prevalence estimates this paper is available online. -

Notice of Non-Compliant Tobacco Product Manufacturers and Brands Removed
Notice of Non-Compliant Tobacco Product Manufacturers and Brands Removed South Carolina has sent Notice of Non-Compliance to the following tobacco product manufacturers as defined in S.C. Code Ann. §11-47-20(i), indicating the intent to remove the company and their brands from the directory as required by S.C. Code Ann. §11-48-30(B)(5): **Removed at the request of the manufacturer. ***Removed because not listed as fire safe compliant. Company Brands Date Removed Kretek International Dreams** June 1, 2020 Commonwealth Brands, Inc. Rave (RYO)** May 15, 2020 Compania Tabacalera Internacional, Director** March 24, 2020 S.A. S&M Brands, Inc. Bailey's** December 31, 2019 Riverside** December 31, 2019 Tahoe** December 31, 2019 Tahoe (RYO)** December 31, 2019 ValuTime** December 31, 2019 Philip Morris USA Inc. Commander** September 13, 2019 English Ovals** September 13, 2019 Native Trading Associates, LLC Native (RYO)** August 15, 2019 Scandinavian Tobacco Group Lane Roll Rich (RYO)** July 15, 2019 Limited Smokin Joes Smokin Joes (RYO)** June 14, 2019 Great Swamp Enterprises Inc Cayuga** May 17, 2019 Seneca Manufacturing Company Heron February 01, 2019 Smokin Joes Smokin Joes January 14, 2019 Smokin Joes Natural January 14, 2019 Cheyenne International LLC Cheyenne RYO** September 01, 2018 Commonwealth Brands, Inc. McClintock** August 15, 2018 King Maker Marketing, Inc. Ace** June 01, 2018 Gold Crest** June 01, 2018 Hi-Val** June 01, 2018 Six Nations Manufacturing Buffalo** May 15, 2018 Gator** May 15, 2018 Stallion** May 15, 2018 NATIONAL TOBACCO Zig Zag (RYO)** April 01, 2018 COMPANY LP ZIG-ZAG** April 01, 2018 Sherman's 1400 Broadway NYC, Naturals** March 01, 2018 Ltd. -

Rotational Health Warnings for Cigarettes File No
United States of America FEDERAL TRADE COMMISSION Washington, D.C. 20580 Division of Advertising Practices MEMORANDUM TO: Public Records Office of the Secretary FROM: Sallie Schools Division of Advertising Practices DATE: May 10, 2013 SUBJECT: Rotational Health Warnings for Cigarettes File No. P854505 Please place the attached documents on the public record in the above-captioned matter. 1. January 19, 2012 letter from Barry M. Boren on behalf of Canadian Agricultural Depot, LLC to Mary Engle. 2. January 23, 2012 letter from Mary K. Engle to Barry M. Boren on behalf of Canadian Agricultural Depot, LLC. 3. January 23, 2012 letter from Nancyellen Keane on behalf of Tisha A. Thompson d/b/a Jacobs Tobacco Company to Mary K. Engle. 4. January 25, 2012 letter from Mary K. Engle to Nancyellen Keane on behalf of Tisha A. Thompson d/b/a Jacobs Tobacco Company. 5. January 18, 2012 letter from William H. Melton, Lignum-2, Inc. to Mary K. Engle. 6. February 6, 2012 letter from Mary K. Engle to William H. Melton, Lignum-2, Inc. 7. February 8, 2012 letter from J. Conrad Seneca, Six Nations Manufacturing to Mary K. Engle. 8. February 9, 2012 letter from Mary K. Engle to J. Conrad Seneca, Six Nations Manufacturing. Public Records May 10, 2013 Page 2 9. February 10, 2012 letter from Henry C. Roemer, III on behalf of Kretek International, Inc. to Sallie Schools. 10. February 13, 2012 letter from Mary K. Engle to Henry C. Roemer, III on behalf of Kretek International, Inc. 11. February 1, 2012 letter from C. -

Tobaccocompaniesbybrand2017 (1)
Geography Category Brand Name (GBO) Company Name (NBO) Data Type 2016 Azerbaijan Cigarettes Viceroy (British American Tobacco Plc) British American Tobacco Azerbaijan MMC Retail Volume 13.0 Azerbaijan Cigarettes Imperial Classic Black (Imperial Brands Plc) Imperial Tobacco MMC Retail Volume 13.0 Azerbaijan Cigarettes LD (Japan Tobacco Inc) JTI Caucasus LLC Retail Volume 8.7 Azerbaijan Cigarettes Winchester (Japan Tobacco Inc) JTI Caucasus LLC Retail Volume 8.6 Azerbaijan Cigarettes Kent (British American Tobacco Plc) British American Tobacco Azerbaijan MMC Retail Volume 7.9 Azerbaijan Cigarettes More (Japan Tobacco Inc) JTI Caucasus LLC Retail Volume 7.5 Azerbaijan Cigarettes Viceroy Lights Blue (British American Tobacco Plc) British American Tobacco Azerbaijan MMC Retail Volume 5.0 Azerbaijan Cigarettes Prima Deluxe (Imperial Brands Plc) Imperial Tobacco MMC Retail Volume 4.4 Azerbaijan Cigarettes West Lights (Imperial Brands Plc) Imperial Tobacco MMC Retail Volume 2.9 Azerbaijan Cigarettes Imperial Classic White (Imperial Brands Plc) Imperial Tobacco MMC Retail Volume 2.4 Azerbaijan Cigarettes Winston (Japan Tobacco Inc) JTI Caucasus LLC Retail Volume 2.0 Azerbaijan Cigarettes Winston Classic (Japan Tobacco Inc) JTI Caucasus LLC Retail Volume 1.7 Azerbaijan Cigarettes Kent Convertibles (British American Tobacco Plc) British American Tobacco Azerbaijan MMC Retail Volume 1.1 Azerbaijan Cigarettes Vogue Superslims Menthe (British American Tobacco Plc) British American Tobacco Azerbaijan MMC Retail Volume 0.9 Azerbaijan Cigarettes Winston