B.GOA.0055 Final Report
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Conservation Management Zones of Australia
Conservation Management Zones of Australia Mitchell Grasslands Prepared by the Department of the Environment Acknowledgements This project and its associated products are the result of collaboration between the Department of the Environment’s Biodiversity Conservation Division and the Environmental Resources Information Network (ERIN). Invaluable input, advice and support were provided by staff and leading researchers from across the Department of Environment (DotE), Department of Agriculture (DoA), the Commonwealth Scientific and Industrial Research Organisation (CSIRO) and the academic community. We would particularly like to thank staff within the Wildlife, Heritage and Marine Division, Parks Australia and the Environment Assessment and Compliance Division of DotE; Nyree Stenekes and Robert Kancans (DoA), Sue McIntyre (CSIRO), Richard Hobbs (University of Western Australia), Michael Hutchinson (ANU); David Lindenmayer and Emma Burns (ANU); and Gilly Llewellyn, Martin Taylor and other staff from the World Wildlife Fund for their generosity and advice. Special thanks to CSIRO researchers Kristen Williams and Simon Ferrier whose modelling of biodiversity patterns underpinned identification of the Conservation Management Zones of Australia. Image Credits Front Cover: Lawn Hill National Park – Peter Lik Page 4: Kowaris (Dasyuroides byrnei) – Leong Lim Page 10: Oriental Pratincole (Glareola maldivarum) – JJ Harrison Page 16: Australian Fossil Mammal Sites (Riversleigh) – World Heritage Listed site – Colin Totterdell Page 18: Mitchell Grasslands -
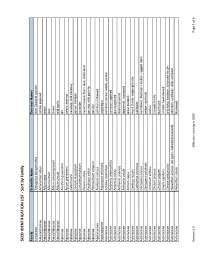
SEED IDENTIFICATION LIST - Sort by Family
SEED IDENTIFICATION LIST - Sort by Family Family Scientific Name Common Names Aizoaceae Tetragonia tetragonoides New Zealand spinach Amaranthaceae Amaranthus albus tumble pigweed Amaryllidaceae Allium cepa onion Amaryllidaceae Allium porrum leek Amaryllidaceae Allium schoenoprasum chives Amaryllidaceae Allium vineale wild garlic Apiaceae Anethum graveolens dill Apiaceae Apium graveolens celery, celeriac Apiaceae Carum carvi caraway; wild caraway Apiaceae Conium maculatum poison hemlock Apiaceae Coriandrum sativum coriander Apiaceae Daucus carota carrot; Queen Ane's lace; wild carrot Apiaceae Pastinaca sativa parsnip; wild parsnip Apiaceae Petroselinum crispum parsley Apocynaceae Asclepias syriaca common milkweed Asparagaceae Asparagus officinalis asparagus Asteraceae Achillea millefolium common yarrow, woolly yarrow Asteraceae Ambrosia artemisiifolia common ragweed Asteraceae Ambrosia trifida giant ragweed Asteraceae Anthemis arvensis field chamomile Asteraceae Anthemis cotula dogfennel, mayweed Asteraceae Arctium lappa great burdock Asteraceae Carduus nutans musk thistle, nodding thistle Asteraceae Carthamus tinctorius safflower Asteraceae Centaurea cyanus cornflower, bachelor's button, ragged robin Asteraceae Centaurea solstitialis yellow starthistle Asteraceae Cichorium endivia endive Asteraceae Cirsium arvense Canada thistle Asteraceae Cirsium vulgare bull thistle Asteraceae Crepis capillaris smooth hawksbeard Asteraceae Cynara cardunculus artichoke, cardoon, artichoke thistle Asteraceae Helianthus annuus (all types, cultivated and -

Complaints Handling Polict
Agnote No: E46 December 2005 Some Important Forage Plants of the Alice Springs District C. Allan, Pastoral Production, Alice Springs The Alice Springs district extends from the South Australian border north to Tennant Creek and east-west from the Queensland border to the Western Australian border. The area occupied by pastoral properties covers some 300,000 km2. This Agnote describes some of the important forage plants in this region. An associated Agnote "Rangeland Pastures of the Alice Springs District" (G4) describes the major plant communities of the region. Plants growing in the Alice Springs district can be considered in three broad categories: • Woody plants (trees and shrubs). These species browsed by cattle are called topfeed. • Grasses (both annual and perennial species). • Herbage (also called forbs). TREES AND SHRUBS Specific information on the description/recognition, nutritive value and grazing management of topfeeds can be found in the booklet "Fodder Trees and Shrubs of the Northern Territory" (copies available from Technical Publications, Department of Primary Industry, Fisheries and Mines). Some of the important trees and shrubs are: Mulga (Acacia aneura) - useful topfeed on better soil types. Easily killed by fire. Hot fires followed by wet years promote re-establishment. Also increases in wet years in the absence of fire. Witchetty bush (Acacia kempeana) - useful topfeed. Re-sprouts after burning. Can increase in wet years on country north of Alice Springs. Rabbits have impacted on its regeneration on much of the calcareous country south of Alice Springs. Ironwood (Acacia estrophiolata) (not to be confused with the poisonous type in the Top End) useful topfeed when accessible to cattle. -

WRA Species Report
Family: Aizoaceae Taxon: Tetragonia tetragonoides Synonym: Demidovia tetragonoides Pall. (basionym) Common Name New Zealand spinach Tetragonia expansa Murray Questionaire : current 20090513 Assessor: Chuck Chimera Designation: H(Hawai'i) Status: Assessor Approved Data Entry Person: Chuck Chimera WRA Score 7 101 Is the species highly domesticated? y=-3, n=0 n 102 Has the species become naturalized where grown? y=1, n=-1 103 Does the species have weedy races? y=1, n=-1 201 Species suited to tropical or subtropical climate(s) - If island is primarily wet habitat, then (0-low; 1-intermediate; 2- High substitute "wet tropical" for "tropical or subtropical" high) (See Appendix 2) 202 Quality of climate match data (0-low; 1-intermediate; 2- High high) (See Appendix 2) 203 Broad climate suitability (environmental versatility) y=1, n=0 y 204 Native or naturalized in regions with tropical or subtropical climates y=1, n=0 y 205 Does the species have a history of repeated introductions outside its natural range? y=-2, ?=-1, n=0 y 301 Naturalized beyond native range y = 1*multiplier (see y Appendix 2), n= question 205 302 Garden/amenity/disturbance weed n=0, y = 1*multiplier (see y Appendix 2) 303 Agricultural/forestry/horticultural weed n=0, y = 2*multiplier (see n Appendix 2) 304 Environmental weed n=0, y = 2*multiplier (see Appendix 2) 305 Congeneric weed n=0, y = 1*multiplier (see Appendix 2) 401 Produces spines, thorns or burrs y=1, n=0 n 402 Allelopathic y=1, n=0 n 403 Parasitic y=1, n=0 n 404 Unpalatable to grazing animals y=1, n=-1 n 405 Toxic -

Asian Vegetables & Herbs Easy to Grow in Southern California
Florence Nishida lagreengrounds.org March 2020 Asian Vegetables & Herbs Easy to Grow in Southern California Name Planting Need Pot Comments time support ok Legumes Snow Peas (Pisum sativum, var. macrocarpon Cool y/n y Bush and pole varieties Spr/Fall Pea shoots – various Spr/Fall y y Pick young, not tendrils Soya Bean (Glycine max) Spr n n Pull up plant for dried Yard Long Bean (Vigna unguiculata ssp. Sesquipedalis) Warm y y Long harvest, keep picked spr Lab Lab Beans (Lab Lab purpureus, Dolichos lab lab), Warm y/n n Can be a perennial if no Hyacinth Bean spr, frost. Contains cyanogenic summer glycosides, eat young or boil mature beans. Winged Bean/Pea (Tetragonolobus purpureus) Cool n y Fast maturing, pretty Brassicas, Headed Chinese cabbage, Napa cabbage (Brassica rapa var. Late n n Fresh – mild sweet flavor, pekinensis) summer, sev. cultivars fall Pak choi/ Bok choy (Brassica rapa var. chinensis), Spring n y Fast-grower, good for celery cabbage inter-cropping, harvest whole or lvs Rosette pak choi (Brassica chinensis var. narinosa), Mid n y Neat, compact, harvest ‘Tatsoi’, ‘Taisai’ summer, leaves or whole fall Flowering Stalk Brassicas Chinese broccoli (Brassica oleracea var. alboglabra) Late spr- n n Young flowering stems and ‘Gai lan’ fall buds Komatsuna (Brassica rapa var. komatsuna) Mustard Late n n Flavor bet. cabbage and spinach spr-fall mustard; eat whole or leaves Mustard Greens Mizuna (Brassica rapa var. japonica), green, red Fall- n y Mild flavored, pretty, pick varieties spring leaves Japanese Giant Red Mustard (Brassica juncea), Osaka Fall-Sprin n y/n Gorgeous, very spicy-hot Purple Mustard (B. -

Bardi Plants an Annotated List of Plants and Their Use
H.,c H'cst. /lust JIus lH8f), 12 (:J): :317-:359 BanE Plants: An Annotated List of Plants and Their Use by the Bardi Aborigines of Dampierland, in North-western Australia \!o\a Smith and .\rpad C. Kalotast Abstract This paper presents a descriptive list of the plants identified and used by the BarcE .\borigines of the Dampierland Peninsula, north~\q:stern Australia. It is not exhaust~ ive. The information is presented in two wavs. First is an alphabetical list of Bardi names including genera and species, use, collection number and references. Second is a list arranged alphabetically according to botanical genera and species, and including family and Bardi name. Previous ethnographic research in the region, vegetation communities and aspects of seasonality (I) and taxonomy arc des~ cribed in the Introduction. Introduction At the time of European colonisation of the south~west Kimberley in the mid nineteenth century, the Bardi Aborigines occupied the northern tip of the Dam pierland Peninsula. To their east lived the island-dwelling Djawi and to the south, the ~yulnyul. Traditionally, Bardi land ownership was based on identification with a particular named huru, translated as home, earth, ground or country. Forty-six bum have been identified (Robinson 1979: 189), and individually they were owned by members of a family tracing their ownership patrilineally, and known by the bum name. Collectively, the buru fall into four regions with names which are roughly equivalent to directions: South: Olonggong; North-west: Culargon; ~orth: Adiol and East: Baniol (Figure 1). These four directional terms bear a superficial resemblance to mainland subsection kinship patterns, in that people sometimes refer to themselves according to the direction in which their land lies, and indeed 'there are. -

Tetragonia Tetragonoides (Pall.) Kuntze (New Zealand Spinach) Prevents Obesity and Hyperuricemia in High-Fat Diet-Induced Obese Mice
nutrients Article Tetragonia tetragonoides (Pall.) Kuntze (New Zealand Spinach) Prevents Obesity and Hyperuricemia in High-Fat Diet-Induced Obese Mice Young-Sil Lee 1, Seung-Hyung Kim 2 ID , Heung Joo Yuk 1, Geung-Joo Lee 3,* and Dong-Seon Kim 1,* 1 Herbal Medicine Research Division, Korea Institute of Oriental Medicine, 1672 Yuseong-daero, Yuseong-gu, Dajeon 34054, Korea; [email protected] (Y.-S.L.); [email protected] (H.J.Y.) 2 Institute of Traditional Medicine and Bioscience, Daejeon University, 62 Daehak-ro, Dong-gu, Daejeon 34520, Korea; [email protected] 3 Department of Horticulture, Chungnam National University, 99 Daehak-ro, Yuseong-gu, Daejeon 34134, Korea * Correspondence: [email protected] (G.-J.L.); [email protected] (D.-S.K.); Tel.: +82-42-821-5734 (G.-J.L.); +82-42-868-9639 (D.-S.K.) Received: 6 July 2018; Accepted: 10 August 2018; Published: 14 August 2018 Abstract: Tetragonia tetragonoides (Pall.) Kuntze, called New Zealand spinach (NZS), is an edible plant used in salad in Western countries and has been used to treat gastrointestinal diseases in traditional medicine. We examined the anti-obesity and anti-hyperuricemic effects of NZS and the underlying mechanisms in high-fat diet (HFD)-induced obese mice. Mice were fed a normal-fat diet (NFD); high-fat diet (HFD); HFD with 75, 150, or 300 mg/kg NZS extract; or 245 mg/kg Garcinia cambogia (GC) extract. NZS decreased body weight gain, total white adipose tissue (WAT), liver weight, and size of adipocytes and improved hepatic and plasma lipid profiles. With NZS, the plasma levels of the leptin and uric acid were significantly decreased while the levels of the adiponectin were increased. -

A Broad Typology of Dry Rainforests on the Western Slopes of New South Wales
A broad typology of dry rainforests on the western slopes of New South Wales Timothy J. Curran1,2, Peter J. Clarke1, and Jeremy J. Bruhl 1 1 Botany, Centre for Ecology, Evolution and Systematics, University of New England, Armidale NSW 2351, AUSTRALIA. 2 Author for correspondence. Current address: The School for Field Studies, Centre for Rainforest Studies, PO Box 141, Yungaburra, Queensland 4884, AUSTRALIA. [email protected] & [email protected] Abstract: Dry rainforests are those communities that have floristic and structural affinities to mesic rainforests and occur in parts of eastern and northern Australia where rainfall is comparatively low and often highly seasonal. The dry rainforests of the western slopes of New South Wales are poorly-understood compared to other dry rainforests in Australia, due to a lack of regional scale studies. This paper attempts to redress this by deriving a broad floristic and structural typology for this vegetation type. Phytogeographical analysis followed full floristic surveys conducted on 400 m2 plots located within dry rainforest across the western slopes of NSW. Cluster analysis and ordination of 208 plots identified six floristic groups. Unlike in some other regional studies of dry rainforest these groups were readily assigned to Webb structural types, based on leaf size classes, leaf retention classes and canopy height. Five community types were described using both floristic and structural data: 1)Ficus rubiginosa–Notelaea microcarpa notophyll vine thicket, 2) Ficus rubiginosa–Alectryon subcinereus–Notelaea microcarpa notophyll vine forest, 3) Elaeodendron australe–Notelaea microcarpa–Geijera parviflora notophyll vine thicket, 4) Notelaea microcarpa– Geijera parviflora–Ehretia membranifolia semi-evergreen vine thicket, and 5) Cadellia pentastylis low microphyll vine forest. -

From Tetragonia Tetragonoides in Lps-Induced Raw 264.7 Cells
EXCLI Journal 2017;16:521-530 – ISSN 1611-2156 Received: January 05, 2017, accepted: March 27, 2017, published: April 18, 2017 Original article: ANTI-INFLAMMATORY ACTIVITY OF HYDROSOLS FROM TETRAGONIA TETRAGONOIDES IN LPS-INDUCED RAW 264.7 CELLS Eun-Yi Koa,1, Su-Hyeon Choa,b,1, Kyungpil Kangc, Gibeom Kimc, Ji-Hyeok Leeb, You-Jin Jeonb, Daekyung Kima, Ginnae Ahnd, Kil-Nam Kime* a Jeju Center, Korea Basic Science Institute (KBSI), Jeju 690-140, Republic of Korea b Department of Marine Life Science, Jeju National University, Jeju 690-756, Republic of Korea c BKSU Lnc. Jeju , 63243, Republic of Korea d Department of Marine Bio-food science, College of Fisheries and Ocean Sciences, Chonnam National University, 500-749, Republic of Korea e Chuncheon Center, Korea Basic Science Institute (KBSI), Chuncheon 24341, Republic of Korea 1 These authors contributed equally to this work. * corresponding author: Tel.: +82-33-815-4607; Fax: +82-33-255-7273; E-mail: [email protected] (KN Kim) http://dx.doi.org/10.17179/excli2017-121 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/). ABSTRACT The present study was performed to investigate the anti-inflammatory activity of Tetragonia tetragonoides hydro- sols (TTH) and its underlying mechanism in lipopolysaccharide (LPS)-induced RAW 264.7 cells. Gas chromatog- raphy (GC) coupled with mass spectrometry and retention index calculations showed that TTH were mainly com- posed of tetratetracontane (29.5 %), nonacosane (27.6 %), and oleamide (17.1 %). TTH significantly decreased the production of nitric oxide (NO), prostaglandin E2 (PGE2), interleukin (IL)-6, and IL-1β in LPS-stimulated RAW 264.7 cells. -

The Palatability, and Potential Toxicity of Australian Weeds to Goats
1 The palatability, and potential toxicity of Australian weeds to goats Helen Simmonds, Peter Holst, Chris Bourke 2 Rural Industries Research and Development Corporation Level 1, AMA House 42 Macquarie Street BARTON ACT 2600 PO Box 4776 KINGSTON ACT 2604 AUSTRALIA © 2000 Rural Industries Research and Development Corporation All rights reserved. The views expressed and the conclusions reached in this publication are those of the authors and not necessarily those of persons consulted. RIRDC shall not be responsible in any way whatsoever to any person who relies in whole or in part on the contents of this report. National Library of Australia Cataloguing in Publication entry: Simmonds, Helen. The palatability, and potential toxicity of Australian weeds to goats. New ed. Includes index. ISBN 0 7347 1216 2 1. Weeds-goats-toxicity-palatability-Australia. i. Holst, Peter. ii. Bourke, Chris. iii. Title. 3 CONTENTS Page Preface i A comment on weed control ii The potential toxicity of weeds to goats.....................................1 Weeds - thought to be highly or moderately toxic to goats ........5 - thought to have low toxicity to goats..........................90 The palatability of weeds to goats..........................................138 The botanical name for weeds listed by their common name 142 The common name for some Australian weeds .....................147 Table of herbicide groups ......................................................151 Further reading .......................................................................152 -

SPECIES REPORT for ASHY STORM-PETREL (Oceanodroma Homochroa)
1 SPECIES REPORT FOR ASHY STORM-PETREL (Oceanodroma homochroa) PHOTO CREDIT: ILANA NIMZ U.S. Fish and Wildlife Service (Service) Bay-Delta Fish and Wildlife Office, Sacramento, California 9/16/13 2 EXECUTIVE SUMMARY The ashy storm-petrel (Oceanodroma homochroa) is a small seabird whose known at-sea distribution ranges from about the California-Oregon Border to Islas San Benitos, Mexico. The 32 known breeding sites of the ashy storm-petrel stretch from Point Cabrillo, Mendocino County, California to Islas Todos Santos Island, Ensenada, Mexico. More than 90 percent of the population breeds in two population centers at Southeast (SE) Farallon Island and in the California Channel Islands. Anacapa, San Miguel, Santa Cruz, Santa Rosa, San Clemente, San Nicholas, Santa Barbara, and Santa Catalina Islands comprise the Channel Islands. Ashy storm-petrels occur at their breeding colonies nearly year-round and occur in greater numbers from February through October. The ashy storm-petrel feeds at night on euphausiids, other krill, decapods, larval lanternfish, fish eggs, young squid, and spiny lobster. Previous Federal Actions The purpose of this species report is to provide the best available scientific and commercial information about the species so that we can evaluate whether or not the species warrants protection under the Endangered Species Act of 1973 (Act or ESA). On August 9, 2009, the Service announced its 12-month finding that found, after reviewing the best available scientific and commercial information, listing the ashy storm-petrel was not warranted. The Center for Biological Diversity challenged this decision in the District Court of the Northern District of California on October 25, 2010. -

A MESSAGE from TRUSTEE HARRY with No 4Wds
Number 87 May 2016 A MESSAGE FROM TRUSTEE HARRY with no 4WDs . If you are planning a private trip prior to or after the meeting Avis & Budget both have an office at Charleville. Prior bookings would be advisable and the cost Greetings Wood Collectors from Staghorn Flat, of 4WDs start at about $150 per day with reductions for longer periods. Esther & I have just returned from the Victorian meeting held on Early booking of Charleville accommodation is essential. Anzac weekend in the Otway’s, staying at Colac as we had never An accommodation guide was printed in the February edi- been in this area before. The Myrtle Beech, Backwoods and tion of DownUnder and is also on the IWCS website as is Euc, regnans were truly remarkable. A report of this meeting the registration form. will feature in DownUnder and on the website. As we travelled home, Esther and I went further west to Planning has continued for the Charleville 2016 International & Quilpie, Eromanga, and then on to the Dig Tree & In- Australasian AGM. We have held meetings with the events coor- namincka, finally passing through Hungerford & Bourke dinator and other key personnel and have also visited Augathella until we reached home. A report on the trees and other and Maryvale Station to complete the final arrangements. A features of our trip are included in this newsletter. copy of the Draft program is included in this edition of 2017 AGM and Conference - Graham Trost has indicted his DownUnder. willingness to coordinate this event in the North Island of The conference registration fee has been set at $475 and the NZ.