Habitability of Planets Around Red Dwarf Stars
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
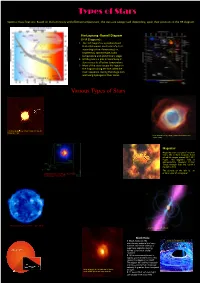
Brown Dwarf: White Dwarf: Hertzsprung -Russell Diagram (H-R
Types of Stars Spectral Classifications: Based on the luminosity and effective temperature , the stars are categorized depending upon their positions in the HR diagram. Hertzsprung -Russell Diagram (H-R Diagram) : 1. The H-R Diagram is a graphical tool that astronomers use to classify stars according to their luminosity (i.e. brightness), spectral type, color, temperature and evolutionary stage. 2. HR diagram is a plot of luminosity of stars versus its effective temperature. 3. Most of the stars occupy the region in the diagram along the line called the main sequence. During that stage stars are fusing hydrogen in their cores. Various Types of Stars Brown Dwarf: White Dwarf: Brown dwarfs are sub-stellar objects After a star like the sun exhausts its nuclear that are not massive enough to sustain fuel, it loses its outer layer as a "planetary nuclear fusion processes. nebula" and leaves behind the remnant "white Since, comparatively they are very cold dwarf" core. objects, it is difficult to detect them. Stars with initial masses Now there are ongoing efforts to study M < 8Msun will end as white dwarfs. them in infrared wavelengths. A typical white dwarf is about the size of the This picture shows a brown dwarf around Earth. a star HD3651 located 36Ly away in It is very dense and hot. A spoonful of white constellation of Pisces. dwarf material on Earth would weigh as much as First directly detected Brown Dwarf HD 3651B. few tons. Image by: ESO The image is of Helix nebula towards constellation of Aquarius hosts a White Dwarf Helix Nebula 6500Ly away. -

The Potential of Planets Orbiting Red Dwarf Stars to Support Oxygenic Photosynthesis and Complex Life
1 The Potential of Planets Orbiting Red Dwarf Stars to Support Oxygenic Photosynthesis and Complex Life Joseph Gale1 and Amri Wandel2 The Institute of Life Sciences1and The Racach Institute of Physics2, The Hebrew University of Jerusalem, 91904, Israel. [email protected] [email protected] Accepted for publication in the International Journal of Astrobiology Abstract We review and reassess the latest findings on the existence of extra-solar system planets and their potential of having environmental conditions that could support Earth-like life. Within the last two decades, the multi-millennial question of the existence of extra-Solar- system planets has been resolved with the discovery of numerous planets orbiting nearby stars, many of which are Earth-sized (and some even with moderate surface temperatures). Of particular interest in our search for clement conditions for extra-solar system life are planets orbiting Red Dwarf (RD) stars, the most numerous stellar type in the Milky Way galaxy. We show that including RDs as potential life supporting host stars could increase the probability of finding biotic planets by a factor of up to a thousand, and reduce the estimate of the distance to our nearest biotic neighbor by up to 10. We argue that multiple star systems need to be taken into account when discussing habitability and the abundance of biotic exoplanets, in particular binaries of which one or both members are RDs. Early considerations indicated that conditions on RD planets would be inimical to life, as their Habitable Zones (where liquid water could exist) would be so close as to make planets tidally locked to their star. -

Educator's Guide: Orion
Legends of the Night Sky Orion Educator’s Guide Grades K - 8 Written By: Dr. Phil Wymer, Ph.D. & Art Klinger Legends of the Night Sky: Orion Educator’s Guide Table of Contents Introduction………………………………………………………………....3 Constellations; General Overview……………………………………..4 Orion…………………………………………………………………………..22 Scorpius……………………………………………………………………….36 Canis Major…………………………………………………………………..45 Canis Minor…………………………………………………………………..52 Lesson Plans………………………………………………………………….56 Coloring Book…………………………………………………………………….….57 Hand Angles……………………………………………………………………….…64 Constellation Research..…………………………………………………….……71 When and Where to View Orion…………………………………….……..…77 Angles For Locating Orion..…………………………………………...……….78 Overhead Projector Punch Out of Orion……………………………………82 Where on Earth is: Thrace, Lemnos, and Crete?.............................83 Appendix………………………………………………………………………86 Copyright©2003, Audio Visual Imagineering, Inc. 2 Legends of the Night Sky: Orion Educator’s Guide Introduction It is our belief that “Legends of the Night sky: Orion” is the best multi-grade (K – 8), multi-disciplinary education package on the market today. It consists of a humorous 24-minute show and educator’s package. The Orion Educator’s Guide is designed for Planetarians, Teachers, and parents. The information is researched, organized, and laid out so that the educator need not spend hours coming up with lesson plans or labs. This has already been accomplished by certified educators. The guide is written to alleviate the fear of space and the night sky (that many elementary and middle school teachers have) when it comes to that section of the science lesson plan. It is an excellent tool that allows the parents to be a part of the learning experience. The guide is devised in such a way that there are plenty of visuals to assist the educator and student in finding the Winter constellations. -

A FEROS Survey of Hot Subdwarf Stars
Open Astron. 2018; 27: 7–13 Research Article Stéphane Vennes*, Péter Németh, and Adela Kawka A FEROS Survey of Hot Subdwarf Stars https://doi.org/10.1515/astro-2018-0005 Received Oct 02, 2017; accepted Nov 07, 2017 Abstract: We have completed a survey of twenty-two ultraviolet-selected hot subdwarfs using the Fiber-fed Extended Range Optical Spectrograph (FEROS) and the 2.2-m telescope at La Silla. The sample includes apparently single objects as well as hot subdwarfs paired with a bright, unresolved companion. The sample was extracted from our GALEX cat- alogue of hot subdwarf stars. We identified three new short-period systems (P = 3.5 hours to 5 days) and determined the orbital parameters of a long-period (P = 62d.66) sdO plus G III system. This particular system should evolve into a close double degenerate system following a second common envelope phase. We also conducted a chemical abundance study of the subdwarfs: Some objects show nitrogen and argon abundance excess with respect to oxygen. We present key results of this programme. Keywords: binaries: close, binaries: spectroscopic, subdwarfs, white dwarfs, ultraviolet: stars 1 Introduction ing a mass transfer event, i.e., comparable to the observed fraction (≈70%) of short period binaries (P < 10 d, Maxted et al. 2001; Morales-Rueda et al. 2003), while the remain- The properties of extreme horizontal branch (EHB) stars, ing single objects are formed by the merger of two helium i.e., the hot, hydrogen-rich (sdB) and helium-rich subd- white dwarfs which would also result in a thin hydrogen warf (sdO) stars, located at the faint blue end of the hori- layer. -

Massive Fast Rotating Highly Magnetized White Dwarfs: Theory and Astrophysical Applications
Massive Fast Rotating Highly Magnetized White Dwarfs: Theory and Astrophysical Applications Thesis Advisors Ph.D. Student Prof. Remo Ruffini Diego Leonardo Caceres Uribe* Dr. Jorge A. Rueda *D.L.C.U. acknowledges the financial support by the International Relativistic Astrophysics (IRAP) Ph.D. program. Academic Year 2016–2017 2 Contents General introduction 4 1 Anomalous X-ray pulsars and Soft Gamma-ray repeaters: A new class of pulsars 9 2 Structure and Stability of non-magnetic White Dwarfs 21 2.1 Introduction . 21 2.2 Structure and Stability of non-rotating non-magnetic white dwarfs 23 2.2.1 Inverse b-decay . 29 2.2.2 General Relativity instability . 31 2.2.3 Mass-radius and mass-central density relations . 32 2.3 Uniformly rotating white dwarfs . 37 2.3.1 The Mass-shedding limit . 38 2.3.2 Secular Instability in rotating and general relativistic con- figurations . 38 2.3.3 Pycnonuclear Reactions . 39 2.3.4 Mass-radius and mass-central density relations . 41 3 Magnetic white dwarfs: Stability and observations 47 3.1 Introduction . 47 3.2 Observations of magnetic white dwarfs . 49 3.2.1 Introduction . 49 3.2.2 Historical background . 51 3.2.3 Mass distribution of magnetic white dwarfs . 53 3.2.4 Spin periods of isolated magnetic white dwarfs . 53 3.2.5 The origin of the magnetic field . 55 3.2.6 Applications . 56 3.2.7 Conclusions . 57 3.3 Stability of Magnetic White Dwarfs . 59 3.3.1 Introduction . 59 3.3.2 Ultra-magnetic white dwarfs . 60 3.3.3 Equation of state and virial theorem violation . -

1. (A) Give the Equation Which Represents Stefan's Law for a Black
PhysicsAndMathsTutor.com 1 1. (a) Give the equation which represents Stefan’s law for a black body source of radiation, defining the symbols used. ..................................................................................................................................... ..................................................................................................................................... ..................................................................................................................................... ..................................................................................................................................... ..................................................................................................................................... (3) (b) (i) The space probe Voyager I is travelling out of the solar system and is, at present, approximately 8000 × 106 km from the Sun. The power source on board is a nuclear generator providing 400 W. If this power had to be obtained from solar panels, calculate the area of the solar panels which would be necessary. power received on Earth from the Sun = 1400 Wm–2 distance of the Earth from the Sun = 150 × 106 km ........................................................................................................................... ........................................................................................................................... .......................................................................................................................... -

February 2018 BRAS Newsletter
Novemb Monthly Meeting Monday, February 12th at 7PM at HRPO er 2017 Issue nd (Monthly meetings are on 2 Mondays, Highland Road Park Observatory) . Program: Star Clusters, a presentation by Rory Bentley. What's In This Issue? President’s Message Secretary's Summary Outreach Report Light Pollution Committee Report Recent Forum Entries 20/20 Vision Campaign Messages from the HRPO Friday Night Lecture Series Globe at Night Adult Astronomy Courses International Astronomy Day Observing Notes – Canis Minor, The Little Dog & Mythology Like this newsletter? See past issues back to 2009 at http://brastro.org/newsletters.html Newsletter of the Baton Rouge Astronomical Society February 2018 © 2018 President’s Message We are now entering the month of February 2018. This month will be unusual for the fact there will be no full moon. This lack of a full moon can happen because the Moon's synodic orbit around Earth takes longer than the 28 days in February. I would remind you that our monthly meeting is on 12th of February at 7 pm. There will be a talk on star clusters given by Rory Bentley. I would also like to remind you of our Business Meeting which will be 7 pm on 7th of February at HRPO. We are investigating ideas which include: an asteroid observing group 2018 Officers: an astrophotography study group President: Steven M. Tilley a BRAS Youtube channel Vice-President: Scott Louque adding additional stargazes for BRAS members Secretary: Krista Reed ways to better utilize BRAS equipment Treasurer: Trey Anding adding another dark sky site BRAS Liaison for BREC: We may not do everything listed if there is not sufficient interest Chris Kersey from members, so if you are willing to help let us know. -
![Arxiv:0908.2624V1 [Astro-Ph.SR] 18 Aug 2009](https://docslib.b-cdn.net/cover/1870/arxiv-0908-2624v1-astro-ph-sr-18-aug-2009-1111870.webp)
Arxiv:0908.2624V1 [Astro-Ph.SR] 18 Aug 2009
Astronomy & Astrophysics Review manuscript No. (will be inserted by the editor) Accurate masses and radii of normal stars: Modern results and applications G. Torres · J. Andersen · A. Gim´enez Received: date / Accepted: date Abstract This paper presents and discusses a critical compilation of accurate, fun- damental determinations of stellar masses and radii. We have identified 95 detached binary systems containing 190 stars (94 eclipsing systems, and α Centauri) that satisfy our criterion that the mass and radius of both stars be known to ±3% or better. All are non-interacting systems, so the stars should have evolved as if they were single. This sample more than doubles that of the earlier similar review by Andersen (1991), extends the mass range at both ends and, for the first time, includes an extragalactic binary. In every case, we have examined the original data and recomputed the stellar parameters with a consistent set of assumptions and physical constants. To these we add interstellar reddening, effective temperature, metal abundance, rotational velocity and apsidal motion determinations when available, and we compute a number of other physical parameters, notably luminosity and distance. These accurate physical parameters reveal the effects of stellar evolution with un- precedented clarity, and we discuss the use of the data in observational tests of stellar evolution models in some detail. Earlier findings of significant structural differences between moderately fast-rotating, mildly active stars and single stars, ascribed to the presence of strong magnetic and spot activity, are confirmed beyond doubt. We also show how the best data can be used to test prescriptions for the subtle interplay be- tween convection, diffusion, and other non-classical effects in stellar models. -

Three Editions of the Star Catalogue of Tycho Brahe*
A&A 516, A28 (2010) Astronomy DOI: 10.1051/0004-6361/201014002 & c ESO 2010 Astrophysics Three editions of the star catalogue of Tycho Brahe Machine-readable versions and comparison with the modern Hipparcos Catalogue F. Verbunt1 andR.H.vanGent2,3 1 Astronomical Institute, Utrecht University, PO Box 80 000, 3508 TA Utrecht, The Netherlands e-mail: [email protected] 2 URU-Explokart, Faculty of Geosciences, Utrecht University, PO Box 80 115, 3508 TC Utrecht, The Netherlands 3 Institute for the History and Foundations of Science, PO Box 80 000, 3508 TA Utrecht, The Netherlands Received 6 January 2010 / Accepted 3 February 2010 ABSTRACT Tycho Brahe completed his catalogue with the positions and magnitudes of 1004 fixed stars in 1598. This catalogue circulated in manuscript form. Brahe edited a shorter version with 777 stars, printed in 1602, and Kepler edited the full catalogue of 1004 stars, printed in 1627. We provide machine-readable versions of the three versions of the catalogue, describe the differences between them and briefly discuss their accuracy on the basis of comparison with modern data from the Hipparcos Catalogue. We also compare our results with earlier analyses by Dreyer (1916, Tychonis Brahe Dani Scripta Astronomica, Vol. II) and Rawlins (1993, DIO, 3, 1), finding good overall agreement. The magnitudes given by Brahe correlate well with modern values, his longitudes and latitudes have error distributions with widths of 2, with excess numbers of stars with larger errors (as compared to Gaussian distributions), in particular for the faintest stars. Errors in positions larger than 10, which comprise about 15% of the entries, are likely due to computing or copying errors. -

Spectropolarimetry
STELLAR PHYSICS WITH HIGH-RESOLUTION UV SPECTROPOLARIMETRY Contact scientist: Julien Morin Laboratoire Univers et Particules de Montpellier (LUPM) Université de Montpellier, CNRS 34095 Montpellier, France [email protected] August 5, 2019 ESA VOYAGE 2050 WHITE PAPER STELLAR PHYSICS WITH HR UV SPECTROPOLARIMETRY ABSTRACT Current burning issues in stellar physics, for both hot and cool stars, concern their magnetism. In hot stars, stable magnetic fields of fossil origin impact their stellar structure and circumstellar environment, with a likely major role in stellar evolution. However, this role is complex and thus poorly understood as of today. It needs to be quantified with high-resolution UV spectropolarimetric measurements. In cool stars, UV spectropolarimetry would provide access to the structure and magnetic field of the very dynamic upper stellar atmosphere, providing key data for new progress to be made on the role of magnetic fields in heating the upper atmospheres, launching stellar winds, and more generally in the interaction of cool stars with their environment (circumstellar disk, planets) along their whole evolution. UV spectropolarimetry is proposed on missions of various sizes and scopes, from POLLUX on the 15-m telescope LUVOIR to the Arago M-size mission dedicated to UV spectropolarimetry. 1 Scientific Context Stars form from material in the interstellar medium (ISM). As they accrete matter from their parent molecular cloud, planets can also form. During the formation and throughout the entire life of stars and planets, a few key basic physical processes, involving in particular magnetic fields, winds, rotation, and binarity, directly affect the internal structure of stars, their dynamics, and immediate circumstellar environment. -

Ultra-Wideband Detection of 22 Coherent Radio Bursts on M Dwarfs
DRAFT VERSION OCTOBER 2, 2018 Typeset using LATEX twocolumn style in AASTeX61 ULTRA-WIDEBAND DETECTION OF 22 COHERENT RADIO BURSTS ON M DWARFS ∗ JACKIE VILLADSEN1, 2 , AND GREGG HALLINAN2 1National Radio Astronomy Observatory 520 Edgemont Rd Charlottesville, VA 22903 2California Institute of Technology 1200 E California Blvd, MC 249-17 Pasadena, CA 91125 ABSTRACT Coherent radio bursts detected from M dwarfs have some analogy with solar radio bursts, but reach orders of magnitude higher luminosities. These events trace particle acceleration, powered by magnetic reconnection, shock fronts (such as formed by coronal mass ejections, CMEs), and magnetospheric currents, in some cases offering the only window into these processes in stellar atmospheres. We conducted a 58-hour, ultra-wideband survey for coherent radio bursts on 5 active M dwarfs. We used the Karl G. Jansky Very Large Array (VLA) to observe simultaneously in three frequency bands covering a subset of 224- 482 MHz and 1-6 GHz, achieving the widest fractional bandwidth to date for any observations of stellar radio bursts. We detected 22 bursts across 13 epochs, providing the first large sample of wideband dynamic spectra of stellar coherent radio bursts. The observed bursts have diverse morphology, with durations ranging from seconds to hours, but all share strong (40-100%) circular polarization. No events resemble solar Type II bursts (often associated with CMEs), but we cannot rule out the occurrence of radio-quiet stellar CMEs. The hours-long bursts are all polarized in the sense of the x-mode of the star’s large-scale magnetic field, suggesting they are cyclotron maser emission from electrons accelerated in the large-scale field, analogous to auroral processes on ultracool dwarfs. -

Stars: Interdisciplinary Exploration Unit 3
ANCHORAGE MUSEUM STARS: INTERDISCIPLINARY EXPLORATION UNIT 3. STARS BACKGROUND INFORMATION KEY TERMS The sun is arguably the most important ingredient for life on Earth. Our sun Star: a hot, dense ball of gaseous material that emits light created by nuclear is one of billions upon billions of stars in the universe. Studying these objects fusion reactions can reveal how our universe, stars, and even planets and life on Earth, have evolved over billions of years. In this unit, you will learn about what stars are Nuclear fusion: the process by which multiple nuclei join together to form a made out of and how they are important factories for seeding the universe with heavier nucleus; this process takes place at the center of all stars and releases most elements found on the periodic table. energy in the form of heat and light STUDENTS WILL: Nebula: a giant cloud of gas and dust out in space; often, nebula are both the birthplaces for stars and the remains left after a star dies • Investigate the composition and anatomy of stars • Learn about the life cycle of stars and stellar evolution Protostar: a dense, hot core of gas and dust, not yet hot enough to start • Explore the night sky and discover examples of different types of stars nuclear fusion; this object is almost a star but has not started nuclear fusion MATERIALS ACTIVITIES • Computer or tablet with internet connection This lesson provides four activity sets and explains each of them in detail • Stars Challenge Activity Sheet on the following pages. Activities should be completed in order.