Proceedings of Kermadec Discoveries and Connections
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hikurangi Plateau: Crustal Structure, Rifted Formation, and Gondwana Subduction History
Article Geochemistry 3 Volume 9, Number 7 Geophysics 3 July 2008 Q07004, doi:10.1029/2007GC001855 GeosystemsG G ISSN: 1525-2027 AN ELECTRONIC JOURNAL OF THE EARTH SCIENCES Published by AGU and the Geochemical Society Click Here for Full Article Hikurangi Plateau: Crustal structure, rifted formation, and Gondwana subduction history Bryan Davy Institute of Geological and Nuclear Sciences, P.O. Box 30368, Lower Hutt, New Zealand ([email protected]) Kaj Hoernle IFM-GEOMAR, Wischhofstraße 1-3, D-24148 Kiel, Germany Reinhard Werner Tethys Geoconsulting GmbH, Wischhofstraße 1-3, D-24148 Kiel, Germany [1] Seismic reflection profiles across the Hikurangi Plateau Large Igneous Province and adjacent margins reveal the faulted volcanic basement and overlying Mesozoic-Cenozoic sedimentary units as well as the structure of the paleoconvergent Gondwana margin at the southern plateau limit. The Hikurangi Plateau crust can be traced 50–100 km southward beneath the Chatham Rise where subduction cessation timing and geometry are interpreted to be variable along the margin. A model fit of the Hikurangi Plateau back against the Manihiki Plateau aligns the Manihiki Scarp with the eastern margin of the Rekohu Embayment. Extensional and rotated block faults which formed during the breakup of the combined Manihiki- Hikurangi plateau are interpreted in seismic sections of the Hikurangi Plateau basement. Guyots and ridge- like seamounts which are widely scattered across the Hikurangi Plateau are interpreted to have formed at 99–89 Ma immediately following Hikurangi Plateau jamming of the Gondwana convergent margin at 100 Ma. Volcanism from this period cannot be separately resolved in the seismic reflection data from basement volcanism; hence seamount formation during Manihiki-Hikurangi Plateau emplacement and breakup (125–120 Ma) cannot be ruled out. -

Greenpeace Deep Sea Oil Briefing
May 2012 Out of our depth: Deep-sea oil exploration in New Zealand greenpeace.org.nz Contents A sea change in Government strategy ......... 4 Safety concerns .............................................. 5 The risks of deep-sea oil ............................... 6 International oil companies in the dock ..... 10 Where is deep-sea oil exploration taking place in New Zealand? ..................... 12 Cover: A view from an altitude of 3200 ft of the oil on the sea surface, originated by the leaking of the Deepwater Horizon wellhead disaster. The BP leased oil platform exploded April 20 and sank after burning, leaking an estimate of more than 200,000 gallons of crude oil per day from the broken pipeline into the sea. © Daniel Beltrá / Greenpeace Right: A penguin lies in oil spilt from the wreck of the Rena © GEMZ Photography 2 l Greenpeace Deep-Sea Oil Briefing l May 2012 The inability of the authorities to cope with the effects of the recent oil spill from the Rena cargo ship, despite the best efforts of Maritime New Zealand, has brought into sharp focus the environmental risks involved in the Government’s decision to open up vast swathes of the country’s coastal waters for deep-sea oil drilling. The Rena accident highlighted the devastation that can be caused by what in global terms is actually still a relatively small oil spill at 350 tonnes and shows the difficulties of mounting a clean-up operation even when the source of the leaking oil is so close to shore. It raised the spectre of the environmental catastrophe that could occur if an accident on the scale of the Deepwater Horizon disaster in the Gulf of Mexico were to occur in New Zealand’s remote waters. -

Quantitative Composition and Granulometry of Aeolian Bedforms in Endeavour and Gale Craters Inferred from Visible Near-Infrared Spectra
45th Lunar and Planetary Science Conference (2014) 1431.pdf QUANTITATIVE COMPOSITION AND GRANULOMETRY OF AEOLIAN BEDFORMS IN ENDEAVOUR AND GALE CRATERS INFERRED FROM VISIBLE NEAR-INFRARED SPECTRA. Mathieu G.A. Lapotre1, Bethany L. Ehlmann1,2, Raymond E. Arvidson3. 1Division of Geological and Planetary Sciences, California Institute of Technology, Pasadena, CA, USA. 2Jet Propulsion Laboratory, California Institute of Technology, Pasadena, CA, USA, 3Department of Earth & Planetary Sciences, Washington University in St. Louis, MO, USA. Introduction: Modern Mars is a wind world. Its ing Spectrometer for Mars (CRISM) visible near- surface hosts a variety of aeolian features, such as line- infrared spectra (VISIR). The goal of this study is to ar, barchan and star dunes, ripples, granule ripples, compare inversions made from orbit to ground truth yardangs and ventifacts [1]. Even though active sand provided by instruments aboard Opportunity at En- transport was observed at the surface [2], it is not clear deavour Crater, Terra Meridiani and Curiosity in Gale whether all of the preserved aeolian bedforms are ac- crater. tive. In particular, transverse aeolian ridges have been We use Hapke’s bidirectional reflectance spectros- suggested to be remnant dunes that formed under past copy theory [6] to invert for optical constants of miner- climatic conditions [3]. als from laboratory spectra [e.g., 7, 8]. These are used Sand transport is largely controlled by the size and to compute single scattering albedos of mineral the density of the grains [4]. Moreover, dunes and rip- endmember components of varying grain sizes. We use ples form in unimodally distributed sand particles from an atmospheric radiative transfer approach, DISORT different instabilities, and the wavelengths of these [9], to correct the CRISM spectra for the effects of the different bedforms do not have the same dependence Martian atmosphere. -

Tailoring of the Controlling and Monitoring Tools for Operations in Shallow Coastal Waters
Tailoring of the controlling and monitoring tools for operations in shallow coastal waters Ref.: D6.1_V1.1 Date: 08/01/2021 Euro-Argo Research Infrastructure Sustainability and Enhancement Project (EA RISE Project) - 824131 Under EC_review This project has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement no 824131. Call INFRADEV-03-2018-2019: Individual support to ESFRI and other world-class research infrastructures Disclaimer: This Deliverable reflects only the author’s views and the European Commission is not responsible for any use that may be made of the information contained therein. 2 Project Euro-Argo RISE - 824131 Deliverable number D6.1 Deliverable title Tailoring of the controlling and monitoring tools for operations in shallow coastal waters Description Report that details the existing Euro-Argo controlling and monitoring tools implemented, developed, tested and tailored for Argo float operations in shallow coastal areas of the Mediterranean, Black and Baltic Seas. Work Package number 6 Work Package title Extension to Marginal Seas Lead Institute OGS Lead authors Giulio Notarstefano Contributors Massimo Pacciaroni, Dimitris Kassis, Atanas Palazov, Violeta Slabakova, Laura Tuomi, Siiriä Simo-Matti, Waldemar Walczowski, Małgorzata Merchel, John Allen, Inmaculada Ruiz, Lara Diaz, Vincent Taillandier, Luca Arduini Plaisant, Romain Cancouët Submission date 08/01/2021 Due date [M18] 30 June 2020 Comments This deliverable was postponed due to Covid-19 pandemic (Article -

Checklist of Fish and Invertebrates Listed in the CITES Appendices
JOINTS NATURE \=^ CONSERVATION COMMITTEE Checklist of fish and mvertebrates Usted in the CITES appendices JNCC REPORT (SSN0963-«OStl JOINT NATURE CONSERVATION COMMITTEE Report distribution Report Number: No. 238 Contract Number/JNCC project number: F7 1-12-332 Date received: 9 June 1995 Report tide: Checklist of fish and invertebrates listed in the CITES appendices Contract tide: Revised Checklists of CITES species database Contractor: World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge, CB3 ODL Comments: A further fish and invertebrate edition in the Checklist series begun by NCC in 1979, revised and brought up to date with current CITES listings Restrictions: Distribution: JNCC report collection 2 copies Nature Conservancy Council for England, HQ, Library 1 copy Scottish Natural Heritage, HQ, Library 1 copy Countryside Council for Wales, HQ, Library 1 copy A T Smail, Copyright Libraries Agent, 100 Euston Road, London, NWl 2HQ 5 copies British Library, Legal Deposit Office, Boston Spa, Wetherby, West Yorkshire, LS23 7BQ 1 copy Chadwick-Healey Ltd, Cambridge Place, Cambridge, CB2 INR 1 copy BIOSIS UK, Garforth House, 54 Michlegate, York, YOl ILF 1 copy CITES Management and Scientific Authorities of EC Member States total 30 copies CITES Authorities, UK Dependencies total 13 copies CITES Secretariat 5 copies CITES Animals Committee chairman 1 copy European Commission DG Xl/D/2 1 copy World Conservation Monitoring Centre 20 copies TRAFFIC International 5 copies Animal Quarantine Station, Heathrow 1 copy Department of the Environment (GWD) 5 copies Foreign & Commonwealth Office (ESED) 1 copy HM Customs & Excise 3 copies M Bradley Taylor (ACPO) 1 copy ^\(\\ Joint Nature Conservation Committee Report No. -

Planktonic Foraminifera As Oceanographic Proxies: Comparison of Biogeographic Classifications Using Some Southwest Pacific Core-Top Faunas
Hindawi Publishing Corporation ISRN Oceanography Volume 2013, Article ID 508184, 15 pages http://dx.doi.org/10.5402/2013/508184 Review Article Planktonic Foraminifera as Oceanographic Proxies: Comparison of Biogeographic Classifications Using Some Southwest Pacific Core-Top Faunas G. H. Scott GNS Science, 1 Fairway Drive, Lower Hutt 5010, New Zealand Correspondence should be addressed to G. H. Scott; [email protected] Received 29 April 2013; Accepted 17 June 2013 Academic Editors: M. Elskens and M. T. Maldonado Copyright © 2013 G. H. Scott. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The distribution of planktonic foraminifera, as free-floating protists, is largely controlled by hydrography. Their death assemblages in surficial sediments provide proxy data on upper water mass properties for paleoceanography. Techniques for mapping faunal distributions for this purpose are compared in a study of 35 core-top samples that span the Subtropical Front in the Southwest Pacific. Faunas are analyzed by taxon composition, order of dominant taxa, and abundance. Taxon composition (presence-absence data) and dominant taxa (ordinal data) recognize groups of sites that approximate major water mass distributions (cool subtropical water, subantarctic water) and clearly define the location of the Subtropical Front. Quantitative data (relative abundances) more closely reflect the success of taxa in upper water mass niches. This information resolves groups of sites that reflect differences in intrawater mass hydrography. Comparisons suggest that abundance data should provide much better oceanographic resolution globally than the widely used ordinal biogeographic classification that identifies only Tropical, Subtropical Transitional, Subpolar and Polar provinces. -

Checklists of Crustacea Decapoda from the Canary and Cape Verde Islands, with an Assessment of Macaronesian and Cape Verde Biogeographic Marine Ecoregions
Zootaxa 4413 (3): 401–448 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2018 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4413.3.1 http://zoobank.org/urn:lsid:zoobank.org:pub:2DF9255A-7C42-42DA-9F48-2BAA6DCEED7E Checklists of Crustacea Decapoda from the Canary and Cape Verde Islands, with an assessment of Macaronesian and Cape Verde biogeographic marine ecoregions JOSÉ A. GONZÁLEZ University of Las Palmas de Gran Canaria, i-UNAT, Campus de Tafira, 35017 Las Palmas de Gran Canaria, Spain. E-mail: [email protected]. ORCID iD: 0000-0001-8584-6731. Abstract The complete list of Canarian marine decapods (last update by González & Quiles 2003, popular book) currently com- prises 374 species/subspecies, grouped in 198 genera and 82 families; whereas the Cape Verdean marine decapods (now fully listed for the first time) are represented by 343 species/subspecies with 201 genera and 80 families. Due to changing environmental conditions, in the last decades many subtropical/tropical taxa have reached the coasts of the Canary Islands. Comparing the carcinofaunal composition and their biogeographic components between the Canary and Cape Verde ar- chipelagos would aid in: validating the appropriateness in separating both archipelagos into different ecoregions (Spalding et al. 2007), and understanding faunal movements between areas of benthic habitat. The consistency of both ecoregions is here compared and validated by assembling their decapod crustacean checklists, analysing their taxa composition, gath- ering their bathymetric data, and comparing their biogeographic patterns. Four main evidences (i.e. different taxa; diver- gent taxa composition; different composition of biogeographic patterns; different endemicity rates) support that separation, especially in coastal benthic decapods; and these parametres combined would be used as a valuable tool at comparing biotas from oceanic archipelagos. -

Integrating Ecology and Justice: the New Papal Encyclical by Mary Evelyn Tucker and John Grim
Feature Integrating Ecology and Justice: The New Papal Encyclical by Mary Evelyn Tucker and John Grim Mat McDermott Una Terra Una Famiglia Humana, One Earth One Family climate march in Vatican City in June 2015. In Brief In June of 2015, Pope Francis released the first encyclical on ecology. The Pope’s message highlights “integral ecology,” intrinsically linking ecological integrity and social justice. While the encyclical notes the statements of prior Popes and Bishops on the environment, Pope Francis has departed from earlier biblical language describing the domination of nature. Instead, he expresses a broader understanding of the beauty and complexity of nature, on which humans fundamentally depend. With “integral ecology” he underscores this connection of humans to the natural environment. This perspective shifts the climate debate to one of a human change of consciousness and conscience. As such, the encyclical has the potential to bring about a tipping point in the global community regarding the climate debate, not merely among Christians, but to all those attending to this moral call to action. 38 | Solutions | July-August 2015 | www.thesolutionsjournal.org n June 18, 2015 Pope Francis thinking. By drawing on and develop- We can compare Pope Francis’ released Laudato Si, the first ing the work of earlier theologians and thinking to the writing of Pope John encyclical in the history of ethicists, this encyclical makes explicit Paul II, who himself builds on Pope O Rerum the Catholic Church on ecology. An the links between social justice and Leo XIII’s progressive encyclical encyclical is the highest-level teaching eco-justice.1 Novarum on workers’ rights in 1891. -
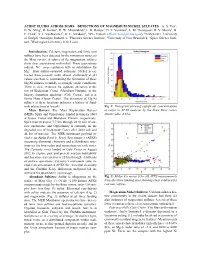
Acidic Fluids Across Mars: Detections of Magnesium-Nickel Sulfates
ACIDIC FLUIDS ACROSS MARS: DETECTIONS OF MAGNESIUM-NICKEL SULFATES. A. S. Yen1, D. W. Ming2, R. Gellert3, D. W. Mittlefehldt2, E. B. Rampe4, D. T. Vaniman5, L. M. Thompson6, R. V. Morris2, B. C. Clark7, S. J. VanBommel3, R. E. Arvidson8, 1JPL- Caltech ([email protected]), 2NASA-JSC, 3University of Guelph, 4Aerodyne Industries, 5Planetary Science Institute, 6University of New Brunswick, 7Space Science Insti- tute, 7Washington University in St. Louis. Introduction: Calcium, magnesium and ferric iron sulfates have been detected by the instrument suites on the Mars rovers. A subset of the magnesium sulfates show clear associations with nickel. These associations indicate Ni2+ co-precipitation with or substitution for Mg2+ from sulfate-saturated solutions. Nickel is ex- tracted from primary rocks almost exclusively at pH values less than 6, constraining the formation of these Mg-Ni sulfates to mildly to strongly acidic conditions. There is clear evidence for aqueous alteration at the rim of Endeavour Crater (Meridiani Planum), in the Murray formation mudstone (Gale Crater), and near Home Plate (Gusev Crater). The discovery of Mg-Ni sulfates at these locations indicates a history of fluid- rock interactions at low pH. Fig 1: Histogram showing significant concentrations Mars Rovers: The Mars Exploration Rovers of sulfur in APXS analyses by the three Mars rovers (MER), Spirit and Opportunity, landed in January 2004 (mean value: 6.6%). at Gusev Crater and Meridiani Planum, respectively. Spirit traversed over 7.7 km through 2210 sols of sur- face operations, and Opportunity is currently on the degraded rim of Endeavour Crater after 4600 sols and 44 km of traverse. -

Vulnerable Marine Ecosystems – Processes and Practices in the High Seas Vulnerable Marine Ecosystems Processes and Practices in the High Seas
ISSN 2070-7010 FAO 595 FISHERIES AND AQUACULTURE TECHNICAL PAPER 595 Vulnerable marine ecosystems – Processes and practices in the high seas Vulnerable marine ecosystems Processes and practices in the high seas This publication, Vulnerable Marine Ecosystems: processes and practices in the high seas, provides regional fisheries management bodies, States, and other interested parties with a summary of existing regional measures to protect vulnerable marine ecosystems from significant adverse impacts caused by deep-sea fisheries using bottom contact gears in the high seas. This publication compiles and summarizes information on the processes and practices of the regional fishery management bodies, with mandates to manage deep-sea fisheries in the high seas, to protect vulnerable marine ecosystems. ISBN 978-92-5-109340-5 ISSN 2070-7010 FAO 9 789251 093405 I5952E/2/03.17 Cover photo credits: Photo descriptions clockwise from top-left: Acanthagorgia spp., Paragorgia arborea, Vase sponges (images courtesy of Fisheries and Oceans, Canada); and Callogorgia spp. (image courtesy of Kirsty Kemp, the Zoological Society of London). FAO FISHERIES AND Vulnerable marine ecosystems AQUACULTURE TECHNICAL Processes and practices in the high seas PAPER 595 Edited by Anthony Thompson FAO Consultant Rome, Italy Jessica Sanders Fisheries Officer FAO Fisheries and Aquaculture Department Rome, Italy Merete Tandstad Fisheries Resources Officer FAO Fisheries and Aquaculture Department Rome, Italy Fabio Carocci Fishery Information Assistant FAO Fisheries and Aquaculture Department Rome, Italy and Jessica Fuller FAO Consultant Rome, Italy FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS Rome, 2016 The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Campbell Plateau: a Major Control on the SW Pacific Sector of The
Ocean Sci. Discuss., doi:10.5194/os-2017-36, 2017 Manuscript under review for journal Ocean Sci. Discussion started: 15 May 2017 c Author(s) 2017. CC-BY 3.0 License. Campbell Plateau: A major control on the SW Pacific sector of the Southern Ocean circulation. Aitana Forcén-Vázquez1,2, Michael J. M. Williams1, Melissa Bowen3, Lionel Carter2, and Helen Bostock1 1NIWA 2Victoria University of Wellington 3The University of Auckland Correspondence to: Aitana ([email protected]) Abstract. New Zealand’s subantarctic region is a dynamic oceanographic zone with the Subtropical Front (STF) to the north and the Subantarctic Front (SAF) to the south. Both the fronts and their associated currents are strongly influenced by topog- raphy: the South Island of New Zealand and the Chatham Rise for the STF, and Macquarie Ridge and Campbell Plateau for the SAF. Here for the first time we present a consistent picture across the subantarctic region of the relationships between front 5 positions, bathymetry and water mass structure using eight high resolution oceanographic sections that span the region. Our results show that the northwest side of Campbell Plateau is comparatively warm due to a southward extension of the STF over the plateau. The SAF is steered south and east by Macquarie Ridge and Campbell Plateau, with waters originating in the SAF also found north of the plateau in the Bounty Trough. Subantarctic Mode Water (SAMW) formation is confirmed to exist south of the plateau on the northern side of the SAF in winter, while on Campbell Plateau a deep reservoir persists into the following 10 autumn. -

THE CORE THEMES of the ENCYCLICAL FRATELLI TUTTI St
THE CORE THEMES OF THE ENCYCLICAL FRATELLI TUTTI St. John Lateran, 15 November 2020 Card. GIANFRANCO RAVASI Preamble Let us begin from a spiritual account drawn from Tibetan Buddhism, a different world from that of the encyclical, but significant nonetheless. A man walks alone on a desert track that dissolves into the distance on the horizon. He suddenly realises that there is another being, difficult to make out, on the same path. It may be a beast that inhabits these deserted spaces; the traveller’s heart beats faster and faster out of fear, as the wilderness does not offer any shelter or help, so he must continue to walk. As he advances, he discerns the profile: a man’s. However, fear does not leave him; in fact, that man could be a fierce bandit. He has no choice but to go on, as the fear of an assault grips his soul. The traveller no longer has the courage to look up. He can hear the other man’s steps approaching. The two men are now face to face: he looks up and stares at the person before him. In his surprise he cries out: “This is my brother whom I haven’t seen for many years!” We wish to refer to this ancient parable from a different religion and culture at the beginning of this presentation to show that the yearning pervading Pope Francis’s new encyclical Fratelli tutti is an essential component of the spiritual breadth of all humanity. In fact, we find a few unexpected “secular” quotations in the text, such as the 1962 album Samba da Bênção by Brazilian poet and musician Vinicius de Moraes (1913-1980) in which he sang: “Life, for all its confrontations, is the art of encounter” (n.