“Inventory of the Meadow Jumping Mouse in Arizona” I09004
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
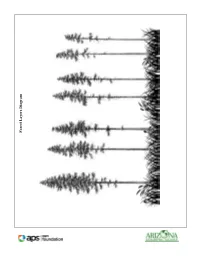
Forest Layers Diagram Forest Layers
Forest Layers Diagram Forest Layers Wildland Fires Background Reading Wildland Fire Wildland fire is a general term describing any non-structure fire that occurs in vegetation and natural fuels. Wildland fire includes both planned and unplanned fires. A prescribed fire is a planned fire intentionally ignited by park managers to meet management objectives. A wildfire is an unplanned fire caused by lightning or other natural causes, by accidental (or arson-caused) human ignitions, or by an escaped prescribed fire. Fuel is one of the ingredients needed for a wildland fire. Fire managers define fuels as all living and dead plant material that can be ignited by a fire. Fuel characteristics strongly influence fire behavior and the resulting fire effects on ecosystems. Types of Fire Spread • Ground fires—which burn organic matter in the soil beneath surface litter and are sustained by glowing combustion. • Surface fires—which spread with a flaming front and burn leaf litter, fallen branches and other fuels located at ground level. • Crown fires—which burn through the top layer of foliage on a tree, known as the canopy or crown. Crown fires are the most intense type of fire and often the most difficult to contain. They may involve strong winds, steep slopes and a heavy fuel load to continue burning. Size and Intensity Even though fire is a fairly common occurrence, very few fires become extremely large. In the United States, 2 to 3 percent of all fires account for more than 95 percent of the total area burned annually. The blackened plots left behind by these types of large fires contribute to the image of fire as a destroyer of the natural environment. -

Land Areas of the National Forest System
United States Department of Agriculture Land Areas of the National Forest System As of September 30, 2018 Forest Service WO Lands FS-383 November 2018 United States Department of Agriculture Forest Service Land Areas of the WO, Lands National Forest FS-383 System November 2018 As of September 30, 2018 Published by: USDA Forest Service 1400 Independence Ave., SW Washington, D.C. 20250-0003 Web site: https://www.fs.fed.us/land/staff/lar-index.shtml Cover photo courtesy of: Chris Chavez Statistics are current as of: 10/15/2018 The National Forest System (NFS) is comprised of: 154 National Forests 58 Purchase Units 20 National Grasslands 7 Land Utilization Projects 17 Research and Experimental Areas 28 Other Areas NFS lands are found in 43 States as well as Puerto Rico and the Virgin Islands. TOTAL NFS ACRES = 192,948,059 NFS lands are organized into: 9 Forest Service Regions 112 Administrative Forest or Forest-level units 506 Ranger District or District-level units The Forest Service administers 128 Wild and Scenic Rivers in 23 States and 446 National Wilderness Areas in 39 States. The FS also administers several other types of nationally-designated areas: 1 National Historic Area in 1 State 1 National Scenic Research Area in 1 State 1 Scenic Recreation Area in 1 State 1 Scenic Wildlife Area in 1 State 2 National Botanical Areas in 1 State 2 National Volcanic Monument Areas in 2 States 2 Recreation Management Areas in 2 States 6 National Protection Areas in 3 States 8 National Scenic Areas in 6 States 12 National Monument Areas in 6 States 12 Special Management Areas in 5 States 21 National Game Refuge or Wildlife Preserves in 12 States 22 National Recreation Areas in 20 States Table of Contents Acreage Calculation ........................................................................................................... -

Biological Opinion for the Revised Programmatic Land Management
United States Department of the Interior U.S. Fish and Wildlife Service Arizona Ecological Services Office 2321 West Royal Palm Road, Suite 103 Phoenix, Arizona 85021-4951 Telephone: (602) 242-0210 FAX: (602) 242-2513 In Reply Refer To: AESO/SE 02EAAZ00-2013-F-0363 May 13, 2015 Mr. Tom Osen, Forest Supervisor Apache-Sitgreaves National Forests Post Office Box 640 Springerville, Arizona 85938-0640 Dear Mr. Osen: Thank you for your May 29, 2014 letter and Biological Assessment (BA), received on that same day, requesting initiation of formal consultation under section 7 of the Endangered Species Act of 1973, as amended (16 U.S.C. 1531 et seq.) (ESA). At issue are impacts that may result from the revised programmatic “Land Management Plan for the Apache Sitgreaves National Forests” (LMP) for lands located in Apache, Navajo, and Greenlee Counties, Arizona (dated January 2013). The proposed action may affect the endangered New Mexico meadow jumping mouse (Zapus hudsonius luteus), the threatened Mexican spotted owl (Strix occidentalis lucida) and its critical habitat, the endangered southwestern willow flycatcher (Empidonax traillii extimus) and its critical habitat, the threatened yellow-billed cuckoo (Coccyzus americanus occidentalis), the threatened northern Mexican gartersnake (Thamnophis eques megalops), the threatened narrow- headed gartersnake (Thamnophis rufipunctatus), the threatened Chiricahua leopard frog (Lithobates chiricahuensis) and its critical habitat, the endangered Three Forks springsnail (Pyrgulopsis trivialis) and its critical habitat, the threatened Apache trout (Oncorhynchus gilae apache), the endangered Gila chub (Gila intermedia) and its critical habitat, the threatened Gila trout (Oncorhynchus gilae gilae), the endangered spikedace (Meda fulgida) and its critical habitat, the endangered loach minnow (Tiaroga cobitis) and its critical habitat, and the threatened Little Colorado spinedace (Lepidomeda vittata) and its critical habitat. -

066 Bear Wallow Wilderness Addition Southeast
Apache-Sitgreaves National Forests Draft Wilderness Evaluation Report Bear Wallow Wilderness Addition Southeast Potential Wilderness PW-03-01-066 June 2009 This draft potential wilderness evaluation report is divided into four parts: background, capability, availability, and need. Capability and availability are intended to be objective evaluations of existing conditions in the Bear Wallow Wilderness Addition Southeast Potential Wilderness. The most important area of focus for public comment is the “need” evaluation. The intent of this part of the evaluation is to consider if the potential wilderness fits into the National Wilderness Preservation System at the regional level. This report offers data that helps us understand different sources that might generate need. Ultimately, however, need for wilderness is generated by public demand. Therefore, public input is an essential component of this part of the potential wilderness evaluation. We would like your feedback on this draft report - if we have missed an important detail or if you would like to share your comments or other input, please contact us. Written comments can be sent to: Apache-Sitgreaves National Forests, Attention: Forest Plan Revision Team, P.O. Box 640, Springerville, Arizona 85938, or E-mail: [email protected]. We are also available by phone; ask for a planning team member at 928-333-4301 or 928-333-6292 (TTY). Table of Contents Introduction..........................................................................................................................................1 -

Allium Gooddingii) Was Selected As One of the Top 20 Species in the Region for Development of Conservation Documents
ACKNOWLEDGMENTS The Goodding’s Onion Conservation Assessment and Strategy and the Goodding’s Onion Cooperative Agreement were developed through the mutual efforts of personnel from the U.S. Forest Service and the U.S. Fish and Wildlife Service. The principal Forest Service participants were Terry Myers, Apache-Sitgreaves National Forests; Mima Parra-Falk, Coronado National Forest; Margaret Kirkeminde, Gila National Forest; Linda Barker, Lincoln National Forest; Heather Hollis, Southwestern Region Regional Office; and Teresa Prendusi, formerly of the Southwestern Region Regional Office. The principal Fish and Wildlife Service participants were Angela Brooks, Arizona Ecological Services Field Office and Charlie McDonald, New Mexico Ecological Services Field Office. Peter Warren of the Arizona Nature Conservancy was helpful during the early stages of strategy development through his in-depth knowledge of Goodding’s onion. Numerous individuals listed in Appendix 3 were helpful in document review and editing. ii CONTENTS ACKNOWLEDGMENTS ................................................ ii CONTENTS ........................................................... iii PART I ) GOODDING’S ONION CONSERVATION ASSESSMENT .............1 I. INTRODUCTION ................................................2 II. REVIEW OF SPECIAL STATUS DESIGNATIONS AND PROTECTIONS . 3 III. NOMENCLATURE AND MORPHOLOGY ...........................5 IV. DISTRIBUTION .................................................6 V. HABITAT AND ECOLOGY .......................................7 -

Grand Canyon Council Oa Where to Go Camping Guide
GRAND CANYON COUNCIL OA WHERE TO GO CAMPING GUIDE GRAND CANYON COUNCIL, BSA OA WHERE TO GO CAMPING GUIDE Table of Contents Introduction to The Order of the Arrow ....................................................................... 1 Wipala Wiki, The Man .................................................................................................. 1 General Information ...................................................................................................... 3 Desert Survival Safety Tips ........................................................................................... 4 Further Information ....................................................................................................... 4 Contact Agencies and Organizations ............................................................................. 5 National Forests ............................................................................................................. 5 U. S. Department Of The Interior - Bureau Of Land Management ................................ 7 Maricopa County Parks And Recreation System: .......................................................... 8 Arizona State Parks: .................................................................................................... 10 National Parks & National Monuments: ...................................................................... 11 Tribal Jurisdictions: ..................................................................................................... 13 On the Road: National -

Page 1464 TITLE 16—CONSERVATION § 1132
§ 1132 TITLE 16—CONSERVATION Page 1464 Department and agency having jurisdiction of, and reports submitted to Congress regard- thereover immediately before its inclusion in ing pending additions, eliminations, or modi- the National Wilderness Preservation System fications. Maps, legal descriptions, and regula- unless otherwise provided by Act of Congress. tions pertaining to wilderness areas within No appropriation shall be available for the pay- their respective jurisdictions also shall be ment of expenses or salaries for the administra- available to the public in the offices of re- tion of the National Wilderness Preservation gional foresters, national forest supervisors, System as a separate unit nor shall any appro- priations be available for additional personnel and forest rangers. stated as being required solely for the purpose of managing or administering areas solely because (b) Review by Secretary of Agriculture of classi- they are included within the National Wilder- fications as primitive areas; Presidential rec- ness Preservation System. ommendations to Congress; approval of Con- (c) ‘‘Wilderness’’ defined gress; size of primitive areas; Gore Range-Ea- A wilderness, in contrast with those areas gles Nest Primitive Area, Colorado where man and his own works dominate the The Secretary of Agriculture shall, within ten landscape, is hereby recognized as an area where years after September 3, 1964, review, as to its the earth and its community of life are un- suitability or nonsuitability for preservation as trammeled by man, where man himself is a visi- wilderness, each area in the national forests tor who does not remain. An area of wilderness classified on September 3, 1964 by the Secretary is further defined to mean in this chapter an area of undeveloped Federal land retaining its of Agriculture or the Chief of the Forest Service primeval character and influence, without per- as ‘‘primitive’’ and report his findings to the manent improvements or human habitation, President. -

Page 1517 TITLE 16—CONSERVATION § 1131 (Pub. L
Page 1517 TITLE 16—CONSERVATION § 1131 (Pub. L. 88–363, § 10, July 7, 1964, 78 Stat. 301.) Sec. 1132. Extent of System. § 1110. Liability 1133. Use of wilderness areas. 1134. State and private lands within wilderness (a) United States areas. The United States Government shall not be 1135. Gifts, bequests, and contributions. liable for any act or omission of the Commission 1136. Annual reports to Congress. or of any person employed by, or assigned or de- § 1131. National Wilderness Preservation System tailed to, the Commission. (a) Establishment; Congressional declaration of (b) Payment; exemption of property from attach- policy; wilderness areas; administration for ment, execution, etc. public use and enjoyment, protection, preser- Any liability of the Commission shall be met vation, and gathering and dissemination of from funds of the Commission to the extent that information; provisions for designation as it is not covered by insurance, or otherwise. wilderness areas Property belonging to the Commission shall be In order to assure that an increasing popu- exempt from attachment, execution, or other lation, accompanied by expanding settlement process for satisfaction of claims, debts, or judg- and growing mechanization, does not occupy ments. and modify all areas within the United States (c) Individual members of Commission and its possessions, leaving no lands designated No liability of the Commission shall be im- for preservation and protection in their natural puted to any member of the Commission solely condition, it is hereby declared to be the policy on the basis that he occupies the position of of the Congress to secure for the American peo- member of the Commission. -

Bear Wallow Wilderness Addition Southeast Potential Wilderness PW-03-01-066
Apache-Sitgreaves National Forests Wilderness Evaluation Report Bear Wallow Wilderness Addition Southeast Potential Wilderness PW-03-01-066 October 2012 Table of Contents Introduction .......................................................................................................................................... 1 Background .......................................................................................................................................... 2 Capability Evaluation ........................................................................................................................... 4 Availability Evaluation ........................................................................................................................ 6 Need Evaluation ................................................................................................................................... 8 Effects of Recommendations ............................................................................................................. 10 Appendix A: Wilderness Evaluation Process ..................................................................................... 17 Appendix B: Capability Evaluation Worksheet ................................................................................. 20 Appendix C: Availability Evaluation Worksheet ............................................................................... 23 Appendix D: Need Evaluation Worksheet ........................................................................................ -

Bear Wallow Wilderness Addition Northwest Potential Wilderness PW-03-01-064
Apache-Sitgreaves National Forests Wilderness Evaluation Report Bear Wallow Wilderness Addition Northwest Potential Wilderness PW-03-01-064 October 2012 Table of Contents Introduction .......................................................................................................................................... 1 Background .......................................................................................................................................... 2 Capability Evaluation ........................................................................................................................... 4 Availability Evaluation ........................................................................................................................ 6 Need Evaluation ................................................................................................................................... 8 Effects of Recommendations ............................................................................................................. 10 Appendix A: Wilderness Evaluation Process ..................................................................................... 17 Appendix B: Capability Evaluation Worksheet ................................................................................. 20 Appendix C: Availability Evaluation Worksheet ............................................................................... 23 Appendix D: Need Evaluation Worksheet ........................................................................................ -

Wallow Fire 2011 Fisheries Report
Wallow Fire 2011 Large Scale Event Recovery Rapid Assessment Team Fisheries Report Apache-Sitgreaves National Forests Submitted by: __/s/ _Kelly Meyer____________________ Kelly Meyer Fish Specialist Region 1, Pinetop Arizona Game and Fish July 31, 2011 FISHERIES Overview: The Wallow fire was the largest recorded fire in Arizona history and burned over 500,000 acres of the Apache-Sitgreaves National Forest. During containment, the Forest initiated a Burned Area Evaluation and Restoration (BAER) group to identify values at risk and initiate emergency protection of these values. The Forest also recognized that it had a need to evaluate and protect these values over a longer time frame. They asked for a Rapid Assessment Team (RAT) to identify actions still needed for emergency and longer term projects. They asked the RAT team to use the Large Scale Event Recovery (LaSER) process to identify these actions. This report is a Fisheries specialist report that is a sub report of the Wallow Fire Rapid Assessment Team report. This fisheries specialist report briefly discusses the present condition and risks to resources, identifies restoration goals, identifies actions, and estimates cost of the actions by fiscal year. The Wallow Fire started May 29, 2011 within the Bear Wallow Wilderness Area located along the southern boundary of the Alpine Ranger District. The fire quickly spread south; with containment declared five weeks later on July 8, 2011 after the fire had burned 557,000 acres. The majority of the fire occurred on National Forest System lands, covering approximately 504,000 acres on the Forest. Approximately 17 percent of the fire was classified as having high soil burn severity, 14 percent moderate soil burn severity, 47 percent low soil burn severity, and 22 percent within the fire perimeter was unburned (Data as of June 24, 2011 BARC map). -

Page 1480 TITLE 16—CONSERVATION § 1113 (Pub
§ 1113 TITLE 16—CONSERVATION Page 1480 (Pub. L. 88–363, § 13, July 7, 1964, 78 Stat. 301.) ment of expenses or salaries for the administra- tion of the National Wilderness Preservation § 1113. Authorization of appropriations System as a separate unit nor shall any appro- There are hereby authorized to be appro- priations be available for additional personnel priated to the Department of the Interior with- stated as being required solely for the purpose of out fiscal year limitation such sums as may be managing or administering areas solely because necessary for the purposes of this chapter and they are included within the National Wilder- the agreement with the Government of Canada ness Preservation System. signed January 22, 1964, article 11 of which pro- (c) ‘‘Wilderness’’ defined vides that the Governments of the United States A wilderness, in contrast with those areas and Canada shall share equally the costs of de- where man and his own works dominate the veloping and the annual cost of operating and landscape, is hereby recognized as an area where maintaining the Roosevelt Campobello Inter- the earth and its community of life are un- national Park. trammeled by man, where man himself is a visi- (Pub. L. 88–363, § 14, July 7, 1964, 78 Stat. 301.) tor who does not remain. An area of wilderness is further defined to mean in this chapter an CHAPTER 23—NATIONAL WILDERNESS area of undeveloped Federal land retaining its PRESERVATION SYSTEM primeval character and influence, without per- manent improvements or human habitation, Sec. which is protected and managed so as to pre- 1131.