Citrus Aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bright Citron™ Soak
Product Profile BRIGHT CITRON™ SOAK WHAT IT IS An aromatic and refreshing sea salt soak for cleansing skin. WHAT IT DOES Softens and cleanses skin. WHY YOU NEED IT • Provides the first step to cleanse impurities and soften skin for a luxurious manicure or pedicure experience. • Gently cleanses without drying skin. FRAGRANCE FEATURED Pink Grapefruit and Warm Amber. FRAGRANCE PROFILE • Bright citrus top notes. • Soft floral mid notes. CREATIVE SUGGESTIONS • Light woody base notes. • Create a fragrant ceremonial experience by placing the Soak into warm water directly prior to ACTIVE BOTANICALS immersing hands or feet. Kaffir Lime (Citrus Hystrix Leaf Extract): • Add lime or grapefruit slices to enhance the Is known to purify skin and promote radiance. experience. • Use as a foot soak prior to all salon services. Honey: • Excellent retail product for home use in bath tubs. Is known to hydrate skin and help retain moisture. INGREDIENT LISTING FEATURED INGREDIENTS & BENEFITS Maris Sal ((Sea Salt) Sel Marin), Sodium Sesquicarbonate, Sea Salt: Argania Spinosa Kernel Oil, Honey, Citrus Hystrix Leaf Natural salt used for its therapeutic properties. Extract, Aqua ((Water) Eau), Glycerin, Propylene Glycol, Salts increase the flow of water in and out of Hydrated Silica, Parfum (Fragrance), Limonene, Linalool, cells, in essence flushing and cleansing the cells. Hexyl Cinnamal, Citral. This facilitates the absorption of other ingredients into the cells. AVAILABLE SIZES • Retail/Professional: 410 g (14.4 oz) Argan Oil (Argania Spinosa Kernel Oil): • Professional: 3.3 kg (118.8 oz) Is known to nourish dry skin. DIRECTIONS FOR USE • For feet, add a teaspoon to footbath and agitate water to dissolve. -

Citrus Limon) and the Future of Its Cultivation by Using Bio-Fertilizers
Available online freely at www.isisn.org Bioscience Research Print ISSN: 1811-9506 Online ISSN: 2218-3973 Journal by Innovative Scientific Information & Services Network REVIEW ARTICLE BIOSCIENCE RESEARCH, 2021 18(1): 889-898. OPEN ACCESS The importance of lemon (Citrus limon) and the future of its cultivation by using bio-fertilizers Salem Abdelmoaty1, Mohammad Moneruzzaman Khandaker*1, Noor Afiza Badaluddin1, Khamsah Suryati Mohd1, Umar Aliu Abdullah1, Nurul Elyni Mat Shaari1, and Khairil Mahmud2 1School of Agriculture Science & Biotechnology, Faculty of Bioresources and Food Industry, Universiti Sultan Zainal Abidin, Besut Campus, 22200 Besut, Terengganu, Malaysia 2Department of Crop Science, Faculty of Agriculture, Universiti Putra Malaysia, Seri Kembangan 43400, Selangor, Malaysia *Correspondence: [email protected] Received 10-12-2020, Revised: 09-03-2021, Accepted: 10-03-2021 e- Published: 15-03-2021 Lemon trees (Citrus lemon L.) are among the trees scattered in Southeast Asia, especially Malaysia, The importance of lemons has increased in recent times, especially after the spread of the Covid-19 epidemic around the world, as lemon juice rich in vitamin C is fortified for the immunity of the human body. Also, it is an important source of Flavonoids, known as antioxidants, which remove free radicals that damage tissue cells within the body. Bio-fertilizers are products containing multiple genera of microorganisms in large numbers and beneficial to both plants and soil in an integrated manner. In addition bio-fertilizers have proved that increase the nutrient content inside plants, whether in leaves or fruits,which is reflected in the quantity and quality of fruits and also balance the ratio between plant nutrients in the soil. -

Characteristic Volatile Fingerprints and Odor Activity Values in Different
molecules Article Characteristic Volatile Fingerprints and Odor Activity Values in Different Citrus-Tea by HS-GC-IMS and HS-SPME-GC-MS Heting Qi 1,2,3 , Shenghua Ding 1,2,3, Zhaoping Pan 1,2,3, Xiang Li 1,2,3 and Fuhua Fu 1,2,3,* 1 Longping Branch Graduate School, Hunan University, Changsha 410125, China; [email protected] (H.Q.); [email protected] (S.D.); [email protected] (Z.P.); [email protected] (X.L.) 2 Provincial Key Laboratory for Fruits and Vegetables Storage Processing and Quality Safety, Agricultural Product Processing Institute, Hunan Academy of Agricultural Sciences, Changsha 410125, China 3 Hunan Province International Joint Lab on Fruits & Vegetables Processing, Quality and Safety, Changsha 410125, China * Correspondence: [email protected]; Tel.: +86-0731-82873369 Received: 24 November 2020; Accepted: 16 December 2020; Published: 19 December 2020 Abstract: Citrus tea is an emerging tea drink produced from tea and the pericarp of citrus, which consumers have increasingly favored due to its potential health effects and unique flavor. This study aimed to simultaneously combine the characteristic volatile fingerprints with the odor activity values (OAVs) of different citrus teas for the first time by headspace gas chromatography-ion mobility spectrometry (HS-GC-IMS) and headspace solid-phase microextraction-gas chromatography-mass spectrometry (HS-SPME-GC-MS). Results showed that the establishment of a citrus tea flavor fingerprint based on HS-GC-IMS data can provide an effective means for the rapid identification and traceability of different citrus varieties. Moreover, 68 volatile compounds (OAV > 1) were identified by HS-SPME-GC-MS, which reflected the contribution of aroma compounds to the characteristic flavor of samples. -

What to Eat on the Autoimmune Protocol
WHAT TO EAT ON THE AUTOIMMUNE PROTOCOL All the foods listed here are great to include in your It’s time to create an epidemic of - health. And it starts with learning ents that will help regulate your immune system and how to eat more nutrient-dense food. your hormones and provide the building blocks that your body needs to heal. You don’t need to eat all of these foods (it’s okay if snails, frog legs, and crickets aren’t your thing, and it’s okay if you just can’t get kangaroo meat or mizuna), but the idea is both to give Poultry innovative ways to increase variety and nutrient density • chicken • grouse • pigeon by exploring new foods. • dove • guinea hen • quail • duck • ostrich • turkey • emu • partridge (essentially, Red Meat • goose • pheasant any bird) • antelope • deer • mutton • bear • elk • pork • beaver • goat • rabbit • beef • hare • sea lion • • horse • seal • boar • kangaroo • whale • camel • lamb (essentially, • caribou • moose any mammal) Amphibians and Reptiles • crocodile • frog • snake • turtle 1 22 Fish* Shellfish • anchovy • gar • • abalone • limpet • scallop • Arctic char • haddock • salmon • clam • lobster • shrimp • Atlantic • hake • sardine • cockle • mussel • snail croaker • halibut • shad • conch • octopus • squid • barcheek • herring • shark • crab • oyster • whelk goby • John Dory • sheepshead • • periwinkle • bass • king • silverside • • prawn • bonito mackerel • smelt • bream • lamprey • snakehead • brill • ling • snapper • brisling • loach • sole • carp • mackerel • • • mahi mahi • tarpon • cod • marlin • tilapia • common dab • • • conger • minnow • trout • crappie • • tub gurnard • croaker • mullet • tuna • drum • pandora • turbot Other Seafood • eel • perch • walleye • anemone • sea squirt • fera • plaice • whiting • caviar/roe • sea urchin • • pollock • • *See page 387 for Selenium Health Benet Values. -

Tropical Horticulture: Lecture 32 1
Tropical Horticulture: Lecture 32 Lecture 32 Citrus Citrus: Citrus spp., Rutaceae Citrus are subtropical, evergreen plants originating in southeast Asia and the Malay archipelago but the precise origins are obscure. There are about 1600 species in the subfamily Aurantioideae. The tribe Citreae has 13 genera, most of which are graft and cross compatible with the genus Citrus. There are some tropical species (pomelo). All Citrus combined are the most important fruit crop next to grape. 1 Tropical Horticulture: Lecture 32 The common features are a superior ovary on a raised disc, transparent (pellucid) dots on leaves, and the presence of aromatic oils in leaves and fruits. Citrus has increased in importance in the United States with the development of frozen concentrate which is much superior to canned citrus juice. Per-capita consumption in the US is extremely high. Citrus mitis (calamondin), a miniature orange, is widely grown as an ornamental house pot plant. History Citrus is first mentioned in Chinese literature in 2200 BCE. First citrus in Europe seems to have been the citron, a fruit which has religious significance in Jewish festivals. Mentioned in 310 BCE by Theophrastus. Lemons and limes and sour orange may have been mutations of the citron. The Romans grew sour orange and lemons in 50–100 CE; the first mention of sweet orange in Europe was made in 1400. Columbus brought citrus on his second voyage in 1493 and the first plantation started in Haiti. In 1565 the first citrus was brought to the US in Saint Augustine. 2 Tropical Horticulture: Lecture 32 Taxonomy Citrus classification based on morphology of mature fruit (e.g. -

Citrus Trifoliata (Rutaceae): Review of Biology and Distribution in the USA
Nesom, G.L. 2014. Citrus trifoliata (Rutaceae): Review of biology and distribution in the USA. Phytoneuron 2014-46: 1–14. Published 1 May 2014. ISSN 2153 733X CITRUS TRIFOLIATA (RUTACEAE): REVIEW OF BIOLOGY AND DISTRIBUTION IN THE USA GUY L. NESOM 2925 Hartwood Drive Fort Worth, Texas 76109 www.guynesom.com ABSTRACT Citrus trifoliata (aka Poncirus trifoliata , trifoliate orange) has become an aggressive colonizer in the southeastern USA, spreading from plantings as a horticultural novelty and use as a hedge. Its currently known naturalized distribution apparently has resulted from many independent introductions from widely dispersed plantings. Seed set is primarily apomictic and the plants are successful in a variety of habitats, in ruderal habits and disturbed communities as well as in intact natural communities from closed canopy bottomlands to open, upland woods. Trifoliate orange is native to southeastern China and Korea. It was introduced into the USA in the early 1800's but apparently was not widely planted until the late 1800's and early 1900's and was not documented as naturalizing until about 1910. Citrus trifoliata L. (trifoliate orange, hardy orange, Chinese bitter orange, mock orange, winter hardy bitter lemon, Japanese bitter lemon) is a deciduous shrub or small tree relatively common in the southeastern USA. The species is native to eastern Asia and has become naturalized in the USA in many habitats, including ruderal sites as well as intact natural commmunities. It has often been grown as a dense hedge and as a horticultural curiosity because of its green stems and stout green thorns (stipular spines), large, white, fragrant flowers, and often prolific production of persistent, golf-ball sized orange fruits that mature in September and October. -

Antioxidant, Antimicrobial, and Other Biological Properties of Pompia Juice
molecules Article Antioxidant, Antimicrobial, and Other Biological Properties of Pompia Juice 1, , 2, 1, 3 Antonio Barberis * y , Monica Deiana y , Ylenia Spissu y , Emanuela Azara , Angela Fadda 1, Pier Andrea Serra 1,4, Guy D’hallewin 1, Marina Pisano 3 , Gabriele Serreli 2, Germano Orrù 1,5 , Alessandra Scano 5, Daniela Steri 6 and Enrico Sanjust 2,* 1 Institute of Sciences of Food Production, National Research Council, 07100 Sassari, Italy; [email protected] (Y.S.); [email protected] (A.F.); [email protected] (P.A.S.); [email protected] (G.D.); [email protected] (G.O.) 2 Department of Biomedical Sciences, University of Cagliari, 09100 Cagliari, Italy; [email protected] (M.D.); [email protected] (G.S.) 3 Institute of Biomolecular Chemistry, National Research Council, 07100 Sassari, Italy; [email protected] (E.A.); [email protected] (M.P.) 4 Department of Medical, Surgical and Experimental Medicine, University of Sassari, 07100 Sassari, Italy 5 Department of Surgical Sciences, Molecular Biology Service, University of Cagliari, 09100 Cagliari, Italy; [email protected] 6 PPD Pharmaceutical Industries, 37121 Verona, Italy; [email protected] * Correspondence: [email protected] (A.B.); [email protected] (E.S.); Tel.: +39-079-2841710 (A.B.) These authors contributed equally to this work. y Received: 15 June 2020; Accepted: 9 July 2020; Published: 13 July 2020 Abstract: Pompia is a Citrus species belonging to Sardinian endemic biodiversity. Health benefits were attributed to its flavedo rind extracts and essential oils while the juice qualities have never been investigated. In this paper, the antioxidant, antimicrobial, and other biological properties of Pompia juice were studied. -

Organic Acids in the Juice of Acid Lemon and Japanese Acid Citrus by Gas Chromatography
九州大学学術情報リポジトリ Kyushu University Institutional Repository Organic Acids in the Juice of Acid Lemon and Japanese Acid Citrus by Gas Chromatography Widodo, Soesiladi E. Fruit Science Laboratory, Faculty of Agriculture, Kyushu University Shiraishi, Mikio Fruit Science Laboratory, Faculty of Agriculture, Kyushu University Shiraishi, Shinichi Fruit Science Laboratory, Faculty of Agriculture, Kyushu University https://doi.org/10.5109/24091 出版情報:九州大学大学院農学研究院紀要. 40 (1/2), pp.39-44, 1995-12. 九州大学農学部 バージョン: 権利関係: ,J. Fat. Agr., Kyushu IJniv., 40 (l-a), 39-44 (1995) Organic Acids in the Juice of Acid Lemon and Japanese Acid Citrus by Gas Chromatography Soesiladi E. Widodo, Mikio Shiraishi and Shinichi Shiraishi Fruit. Science Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka 81 l-23, Japan (RWC~i/‘~?C~ C/1/?1P 15, 199<5) Acetate, glycolate, butyratc, oxalate, malonate, succinate, furnaratp, glyoxylate, malate, tattarate, cis-aconitatc and citrate were detected in the juice of Hanayu (Ci7tnt.s /ttrr/c~jrr Hart. ex Shirai), Daidai (C:. tr/i,rnt/tGt /II Linn. var. Cynthifera Y. Tanaka), Kabosu (6’. .sp/~rc:r~oc~r ~IXI Hart,. cx Tanaka), ‘Lisbon lemon (C.limon Burm. f. Lisbon) and Yuzu (C:.,jrr/ros Sieb. ex Tanaka) with compositions and contcnt,s varied according t,o sampling years and species. Citrate and rnalat,e were predominant, accounting for more than 90% and 3-9% of the total detected acids, respect.ively. The other acids presented in tracts, accounting t.ol.ally for roughly less than 0.5%. INTRODUCTION A number of chromatographic methods have been employed for determining organic acids (OAs) in citrus extracts. -
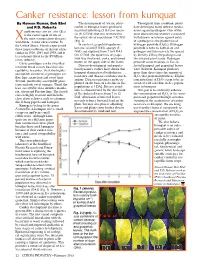
Canker Resistance: Lesson from Kumquat by Naveen Kumar, Bob Ebel the Development of Asiatic Citrus Throughout Their Evolution, Plants and P.D
Canker resistance: lesson from kumquat By Naveen Kumar, Bob Ebel The development of Asiatic citrus Throughout their evolution, plants and P.D. Roberts canker in kumquat leaves produced have developed many defense mecha- anthomonas citri pv. citri (Xcc) localized yellowing (5 DAI) or necro- nisms against pathogens. One of the is the causal agent of one of sis (9-12 DAI) that was restricted to most characteristic features associated the most serious citrus diseases the actual site of inoculation 7-12 DAI with disease resistance against entry X (Fig. 2). of a pathogen is the production of worldwide, Asiatic citrus canker. In the United States, Florida experienced In contrast, grapefruit epidermis hydrogen peroxide (H2O2). Hydrogen three major outbreaks of Asiatic citrus became raised (5 DAI), spongy (5 peroxide is toxic to both plant and canker in 1910, 1984 and 1995, and it DAI) and ruptured from 7 to 8 DAI. pathogen and thus restricts the spread is a constant threat to the $9 billion On 12 DAI, the epidermis of grape- by directly killing the pathogen and citrus industry. fruit was thickened, corky, and turned the infected plant tissue. Hydrogen Citrus genotypes can be classified brown on the upper side of the leaves. peroxide concentrations in Xcc-in- into four broad classes based on sus- Disease development and popula- fected kumquat and grapefruit leaves ceptibility to canker. First, the highly- tion dynamics studies have shown that were different. Kumquat produces susceptible commercial genotypes are kumquat demonstrated both disease more than three times the amount of Key lime, grapefruit and sweet lime. -

Freeze Response of Citrus and Citrus- Speeds (Nisbitt Et Al., 2000)
HORTSCIENCE 49(8):1010–1016. 2014. and tree and grove size (Bourgeois et al., 1990; Ebel et al., 2005). Protection using microsprinklers is compromised by high wind Freeze Response of Citrus and Citrus- speeds (Nisbitt et al., 2000). Developing more cold-tolerant citrus varieties through breeding related Genotypes in a Florida Field and selection has long been considered the most effective long-term solution (Grosser Planting et al., 2000; Yelenosky, 1985). Citrus and Citrus relatives are members Sharon Inch, Ed Stover1, and Randall Driggers of the family Rutaceae. The subtribe Citrinae U.S. Horticultural Research Laboratory, U.S. Department of Agriculture, is composed of Citrus (mandarins, oranges, Agricultural Research Service, 2001 South Rock Road, Fort Pierce, FL pummelos, grapefruits, papedas, limes, lem- ons, citrons, and sour oranges); Poncirus 34945 (deciduous trifoliate oranges); Fortunella Richard F. Lee (kumquats); Microcitrus and Eremocitrus (both Australian natives); and Clymenia National Clonal Germplasm Repository for Citrus and Dates, U.S. (Penjor et al., 2013). There is considerable Department of Agriculture, Agricultural Research Service, 1060 Martin morphological and ecological variation within Luther King Boulevard, Riverside, CA 92521 this group. With Citrus, cold-hardiness ranges from cold-tolerant to cold-sensitive (Soost and Additional index words. Aurantioideae, citrus breeding, cold-sensitive, defoliation, dieback, Roose, 1996). Poncirus and Fortunella are frost damage, Rutaceae, Toddalioideae considered the most cold-tolerant genera that Abstract. A test population consisting of progenies of 92 seed-source genotypes (hereafter are cross-compatible with Citrus. Poncirus called ‘‘parent genotypes’’) of Citrus and Citrus relatives in the field in east–central trifoliata reportedly can withstand tempera- Florida was assessed after natural freeze events in the winters of 2010 and 2011. -

FEMA GRAS Assessment of Natural Flavor Complexes Citrus-Derived
Food and Chemical Toxicology 124 (2019) 192–218 Contents lists available at ScienceDirect Food and Chemical Toxicology journal homepage: www.elsevier.com/locate/foodchemtox FEMA GRAS assessment of natural flavor complexes: Citrus-derived T flavoring ingredients Samuel M. Cohena, Gerhard Eisenbrandb, Shoji Fukushimac, Nigel J. Gooderhamd, F. Peter Guengeriche, Stephen S. Hechtf, Ivonne M.C.M. Rietjensg, Maria Bastakih, ∗ Jeanne M. Davidsenh, Christie L. Harmanh, Margaret McGowenh, Sean V. Taylori, a Havlik-Wall Professor of Oncology, Dept. of Pathology and Microbiology, University of Nebraska Medical Center, 983135 Nebraska Medical Center, Omaha, NE, 68198- 3135, USA b Food Chemistry & Toxicology, Kühler Grund 48/1, 69126 Heidelberg, Germany c Japan Bioassay Research Center, 2445 Hirasawa, Hadano, Kanagawa, 257-0015, Japan d Dept. of Surgery and Cancer, Imperial College London, Sir Alexander Fleming Building, London, SW7 2AZ, United Kingdom e Dept. of Biochemistry, Vanderbilt University School of Medicine, Nashville, TN, 37232-0146, USA f Masonic Cancer Center, Dept. of Laboratory Medicine and Pathology, University of Minnesota, Cancer and Cardiovascular Research Building, 2231 6th St. SE, Minneapolis, MN, 55455, USA g Division of Toxicology, Wageningen University, Stippeneng 4, 6708 WE, Wageningen, the Netherlands h Flavor and Extract Manufacturers Association, 1101 17th Street, NW Suite 700, Washington, DC, 20036, USA i Scientific Secretary to the FEMA Expert Panel, 1101 17th Street, NW Suite 700, Washington, DC,20036,USA ARTICLE INFO ABSTRACT Keywords: In 2015, the Expert Panel of the Flavor and Extract Manufacturers Association (FEMA) initiated a re-evaluation Citrus of the safety of over 250 natural flavor complexes (NFCs) used as flavoring ingredients. This publication isthe Natural flavor complex first in a series and summarizes the evaluation of54 Citrus-derived NFCs using the procedure outlined in Smith Botanical et al. -

Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives
plants Article Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives Clémentine Baccati 1, Marc Gibernau 1, Mathieu Paoli 1 , Patrick Ollitrault 2,3 ,Félix Tomi 1,* and François Luro 2 1 Laboratoire Sciences Pour l’Environnement, Equipe Chimie et Biomasse, Université de Corse—CNRS, UMR 6134 SPE, Route des Sanguinaires, 20000 Ajaccio, France; [email protected] (C.B.); [email protected] (M.G.); [email protected] (M.P.) 2 UMR AGAP Institut, Université Montpellier, CIRAD, INRAE, Institut Agro, 20230 San Giuliano, France; [email protected] (P.O.); [email protected] (F.L.) 3 CIRAD, UMR AGAP, 20230 San Giuliano, France * Correspondence: [email protected]; Tel.: +33-495-52-4122 Abstract: The Papeda Citrus subgenus includes several species belonging to two genetically distinct groups, containing mostly little-exploited wild forms of citrus. However, little is known about the potentially large and novel aromatic diversity contained in these wild citruses. In this study, we characterized and compared the essential oils obtained from peels and leaves from representatives of both Papeda groups, and three related hybrids. Using a combination of GC, GC-MS, and 13C-NMR spectrometry, we identified a total of 60 compounds in peel oils (PO), and 76 compounds in leaf oils (LO). Limonene was the major component in almost all citrus PO, except for C. micrantha and C. hystrix, where β-pinene dominated (around 35%). LO composition was more variable, with different Citation: Baccati, C.; Gibernau, M.; major compounds among almost all samples, except for two citrus pairs: C.