Spatial and Temporal Variation in Atlantic Salmon Abundance in The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ST. MARY's the District of Placentia
PLACENTIA – ST. MARY’S The District of Placentia – St. Mary’s shall consist of and include all that part of the Province of Newfoundland and Labrador bounded as follows: Beginning at the intersection of the eastern shoreline of Chapel Arm, Trinity Bay, and the Town of Chapel Arm Municipal Boundary (2001); Thence running in a general southeasterly direction along the said Municipal Boundary to its intersection with the centre line of the Trans Canada Highway; Thence running in a general northwesterly direction along the centre line of the Trans Canada Highway to its intersection with the Meridian of 53o40’ West Longitude; Thence running due south along the Meridian of 53o40’ West Longitude to its intersection with the Parallel of 47o25’ North Latitude; Thence running due west along the Parallel of 47o25’ North Latitude to its intersection with the Town of Long Harbour-Mount Arlington Heights Municipal Boundary (2010); Thence running in a general southwesterly and northwesterly direction along the said Municipal Boundary to its intersection with the southern shoreline of Long Harbour; Thence running in a general southwesterly direction along the sinuosities of Placentia Bay to a point of land known as Point Lance; Thence running in a general northeasterly and southerly direction along the sinuosities of St. Mary’s Bay to a point of land known as Western Head; Thence running in a northeasterly direction along a line to the intersection of the centre line of the Trans Canada Highway and the centre line of Witless Bay Line, Route 13; Thence -
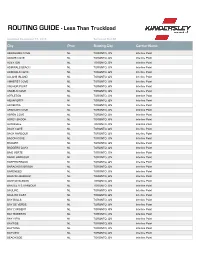
ROUTING GUIDE - Less Than Truckload
ROUTING GUIDE - Less Than Truckload Updated December 17, 2019 Serviced Out Of City Prov Routing City Carrier Name ABRAHAMS COVE NL TORONTO, ON Interline Point ADAMS COVE NL TORONTO, ON Interline Point ADEYTON NL TORONTO, ON Interline Point ADMIRALS BEACH NL TORONTO, ON Interline Point ADMIRALS COVE NL TORONTO, ON Interline Point ALLANS ISLAND NL TORONTO, ON Interline Point AMHERST COVE NL TORONTO, ON Interline Point ANCHOR POINT NL TORONTO, ON Interline Point ANGELS COVE NL TORONTO, ON Interline Point APPLETON NL TORONTO, ON Interline Point AQUAFORTE NL TORONTO, ON Interline Point ARGENTIA NL TORONTO, ON Interline Point ARNOLDS COVE NL TORONTO, ON Interline Point ASPEN COVE NL TORONTO, ON Interline Point ASPEY BROOK NL TORONTO, ON Interline Point AVONDALE NL TORONTO, ON Interline Point BACK COVE NL TORONTO, ON Interline Point BACK HARBOUR NL TORONTO, ON Interline Point BACON COVE NL TORONTO, ON Interline Point BADGER NL TORONTO, ON Interline Point BADGERS QUAY NL TORONTO, ON Interline Point BAIE VERTE NL TORONTO, ON Interline Point BAINE HARBOUR NL TORONTO, ON Interline Point BAKERS BROOK NL TORONTO, ON Interline Point BARACHOIS BROOK NL TORONTO, ON Interline Point BARENEED NL TORONTO, ON Interline Point BARR'D HARBOUR NL TORONTO, ON Interline Point BARR'D ISLANDS NL TORONTO, ON Interline Point BARTLETTS HARBOUR NL TORONTO, ON Interline Point BAULINE NL TORONTO, ON Interline Point BAULINE EAST NL TORONTO, ON Interline Point BAY BULLS NL TORONTO, ON Interline Point BAY DE VERDE NL TORONTO, ON Interline Point BAY L'ARGENT NL TORONTO, ON -

Labrador; These Will Be Done During the Summer
Fisheries Peches I and Oceans et Oceans 0 NEWFOUNDLAND REGION ((ANNUAL REPORT 1985-86 Canada ) ceare SMALL CRAFT HARBOURS BRANCH Y.'• ;'''' . ./ DEPARTMENT OF FISHERIES AND OCEANS NEWFOUNDLAND REGION . 0 4.s.'73 ' ANNUAL REPORT - 1985/86 R edlioft TABLE OF CONTENTS PAGE 1. Overview and Summary 1 2. Small Craft Harbours Branch National Planning Framework 3 3. Long Range Planning: Nfld. Region 8 4. Project Evaluation 10 5. Harbour Maintenance and Development Programs 11 6. Harbour Operations 16 7. Budget Utilization (Summary) 1985/86 17 APPENDICES 1. Photos 2. Harbour Classification 3. Minimum Services Offered 4. Condition Rating Scale 5. Examples of Project Type 6. Project Evaluation 7. Regular Program Projects 1985/86 8. Joint SCH-Job Creation Projects 1984/85/86 9. Joint SCH-Job Creation Projects 1985/86/87 10. Dredging Projects Utilizing DPW Plant 11. Advance Planning 12. Property Acquisition Underway 1 OVERVIEW AND SUMMARY Since the establishment of Small Craft Harbours Branch of the Department of Fisheries and Oceans in 1973, the Branch has been providing facilities such as breakwaters, wharves, slipways, gear storage, shore protection, floats and the dredging of channels and basins, in fishing and recreational harbours within the Newfoundland Region. This third annual report produced by Small Craft Harbours Branch, Newfoundland Region, covers the major activities of the Branch for the fiscal year 1985/86. During the fiscal year continuing efforts were made towards planning of the Small Craft Harbours Program to better define and priorize projects, and to maximize the socio-economic benefits to the commercial fishing industry. This has been an on-going process and additional emphasis was placed on this activity over the past three years. -
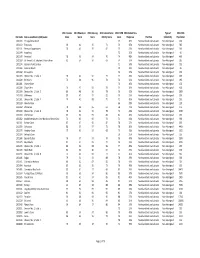
CSD Code Census Subdivision (CSD) Name 2011 Income Score
2011 Income 2011 Education 2011 Housing 2011 Labour Force 2011 CWB 2011 Global Non‐ Type of 2011 NHS CSD Code Census subdivision (CSD) name Score Score Score Activity Score Score Response Province Collectivity Population 1001105 Portugal Cove South 67 36% Newfoundland and Labrador Non‐Aboriginal 160 1001113 Trepassey 90 42 95 71 74 35% Newfoundland and Labrador Non‐Aboriginal 545 1001131 Renews‐Cappahayden 78 46 95 82 75 35% Newfoundland and Labrador Non‐Aboriginal 310 1001144 Aquaforte 72 31% Newfoundland and Labrador Non‐Aboriginal 90 1001149 Ferryland 78 53 94 70 74 48% Newfoundland and Labrador Non‐Aboriginal 465 1001169 St. Vincent's‐St. Stephen's‐Peter's River 81 54 94 69 74 37% Newfoundland and Labrador Non‐Aboriginal 315 1001174 Gaskiers‐Point La Haye 71 39% Newfoundland and Labrador Non‐Aboriginal 235 1001186 Admirals Beach 79 22% Newfoundland and Labrador Non‐Aboriginal 85 1001192 St. Joseph's 72 27% Newfoundland and Labrador Non‐Aboriginal 125 1001203 Division No. 1, Subd. X 76 44 91 77 72 45% Newfoundland and Labrador Non‐Aboriginal 495 1001228 St. Bride's 76 38 96 78 72 24% Newfoundland and Labrador Non‐Aboriginal 295 1001281 Chance Cove 74 40% Newfoundland and Labrador Non‐Aboriginal 120 1001289 Chapel Arm 79 47 92 78 74 38% Newfoundland and Labrador Non‐Aboriginal 405 1001304 Division No. 1, Subd. E 80 48 96 78 76 20% Newfoundland and Labrador Non‐Aboriginal 2990 1001308 Whiteway 80 50 93 82 76 25% Newfoundland and Labrador Non‐Aboriginal 255 1001321 Division No. 1, Subd. F 74 41 98 70 71 45% Newfoundland and Labrador Non‐Aboriginal 550 1001328 New Perlican 66 28% Newfoundland and Labrador Non‐Aboriginal 120 1001332 Winterton 78 38 95 61 68 41% Newfoundland and Labrador Non‐Aboriginal 475 1001339 Division No. -

EIGHTH SUNDAY in ORDINARY TIME March 1, 2020
Good Shepherd Parish EIGHTH SUNDAY P.O. Box 30 St. Joseph’s, NL A0B 3A0 IN ORDINARY TIME Toll Free Number: 1-888-521-2888 March 1, 2020 Telephone Number: 521-2206 Fax Number: 709-521-2888 Pastor: Very Rev. Fr. Stephen J Courtney Archdiocese of St. John’s Toll Free 1-800-563-7700 ext. 250 Or my Direct Line 709-726-9197 Administrative Assistant: Betty Kielly PARISH OFFICE HOURS: Monday - Friday: 9:00 am –12:00 and 1:00pm – 5:00pm Good Shepherd Parish Website www.gsparish.ca: Archdiocese of St. John’s www.rcsj.org Facebook (@GoodShepherdParishSMB) Good Shepherd Parish New Email Address [email protected] PARISH COUNCIL CHAIRPERSON: Shirley Kielly 525-2936 PARISH FINANCE COUNCIL CHAIRPERSON: Howard Power 521-2431 MARRIAGE NOTIFICATIONS: 6 Months (Marriage Preparation course required) Please call Theresa Whiffen (RCEC) to register at 726-3660 ext. 236 PRE-BAPTISM COURSE: To register please call Dave Kenny @ 229-7395 SACRAMENT OF RECONCILIATION available before all Masses, or anytime by appointment at the parish house. VISITATION TO THE SICK AND ANOINTING OF THE SICK Please call the Parish Office if you would like Father Steve to visit you at home or in the hospital. MASS SCHEDULE (When there is a funeral, weekday mass will be re-scheduled) DATE TIME LOCATION MASS INTENTIONS Saturday 4:00 pm Mount Carmel Lori Rohan; Clyde Bennett March 7 Rita Lewis Ministry Server: Ignatius Philpott Readers: Hughie Lawlor, Lucy Hawco Gifts: Marie Gregory & family Eucharistic Minister: Richard Fowler Sunday 10:30 am St. Mary’s Jack & Mary Vail & deceased family members March 8 Simon Tobin & deceased family members Bern Halleran Ministry Cross: Len Ryan Readers: Thelma Stamp, Bill Breen Gifts: Ron & Sylvia Molloy Eucharistic Minister: Helena Critch LENTEN PENITENIAL SERVICE Thursday March 19: 6:30 pm Mount Carmel Wednesday March 25: 6:30 pm St. -
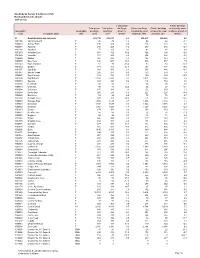
CSD) Newfoundland and Labrador 2016 Census
Dwellings by Census Subdivision (CSD) Newfoundland and Labrador 2016 Census Total private Private dwellings Total private Total private dwellings, Private dwellings Private dwellings occupied by usual Geographic Geographic dwellings, dwellings, growth or occupied by usual occupied by usual residents, growth or code Geographic name type 2016 2011 decline residents, 2016 residents, 2011 decline 10 Newfoundland and Labrador 265,739 250,275 6.2 218,673 208,842 4.7 1001186 Admirals Beach T 80 79 1.3 62 64 -3.1 1009023 Anchor Point T 130 129 0.8 124 121 2.5 1006011 Appleton T 246 265 -7.2 239 246 -2.8 1001144 Aquaforte T 71 65 9.2 41 44 -6.8 1001270 Arnold's Cove T 537 462 16.2 392 400 -2.0 1001464 Avondale T 373 349 6.9 294 282 4.3 1006026 Badger T 325 345 -5.8 302 319 -5.3 1008079 Baie Verte T 646 587 10.1 545 537 1.5 1002040 Baine Harbour T 61 73 -16.4 52 58 -10.3 1001512 Bauline T 174 143 21.7 163 137 19.0 1001557 Bay Bulls T 589 545 8.1 566 486 16.5 1001347 Bay de Verde T 216 207 4.3 173 170 1.8 1002027 Bay L'Argent T 132 143 -7.7 109 129 -15.5 1001409 Bay Roberts T 2,658 2,434 9.2 2,413 2,244 7.5 1008040 Baytona T 129 125 3.2 116 112 3.6 1008066 Beachside T 80 81 -1.2 64 66 -3.0 1009042 Bellburns T 46 41 12.2 26 27 -3.7 1003004 Belleoram T 175 181 -3.3 152 150 1.3 1008038 Birchy Bay T 260 276 -5.8 227 228 -0.4 1009026 Bird Cove T 93 89 4.5 76 74 2.7 1001394 Bishop's Cove T 147 128 14.8 125 110 13.6 1006019 Bishop's Falls T 1,502 1,435 4.7 1,396 1,381 1.1 1007023 Bonavista T 1,749 1,688 3.6 1,520 1,515 0.3 1006022 Botwood T 1,302 1,326 -

Newfoundland & Labrador Paint Stewardship Program
Newfoundland & Labrador Paint Stewardship Program 2017 Annual Report Submitted to: Multi-Materials Stewardship Board Submitted by: Product Care Association of Canada Date: April 30, 2018 Table of Contents 1.0 About Product Care Association of Canada ...................................................................................... 2 2.0 Brand Owner Information ................................................................................................................. 3 3.0 Collection .......................................................................................................................................... 4 4.0 Processing ....................................................................................................................................... 12 5.0 Communication and Education ....................................................................................................... 18 6.0 Financial Information ...................................................................................................................... 20 APPENDIX 1 – Collection Sites as of December 31, 2017 ........................................................................... 21 APPENDIX 2 – Collection Site Locator ......................................................................................................... 23 APPENDIX 3 – PoS and PoR Materials ......................................................................................................... 24 APPENDIX 4 - Sample Facebook Post ......................................................................................................... -
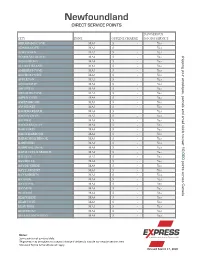
Newfoundland DIRECT SERVICE POINTS
Newfoundland DIRECT SERVICE POINTS DANGEROUS CITY ZONE OFFLINE CHARGE GOODS SERVICE ABRAHAMS COVE MA4 $ - Yes ADAMS COVE MA3 $ - Yes ADEYTOWN MA3 $ - Yes ADMIRALS BEACH MA4 $ - Yes shipping your envelopes, parcels and small skids to over AGUATHUNA MA4 $ - Yes ALLANS ISLAND MA3 $ - Yes AMHERST COVE MA3 $ - Yes ANCHOR POINT MA4 $ - Yes APPLETON MA3 $ - Yes AQUAFORTE MA4 $ - Yes ARGENTIA MA4 $ - Yes ARNOLDS COVE MA3 $ - Yes ASPEN COVE MA4 $ - Yes ASPEY BROOK MA3 $ - Yes AVONDALE MA3 $ - Yes BACK HARBOUR MA4 $ - Yes BACON COVE MA3 $ - Yes BADGER MA4 $ - Yes BADGERS QUAY MA4 $ - Yes BAIE VERTE MA4 $ - Yes BAINE HARBOUR MA4 $ - Yes BARACHOIS BROOK MA4 $ - Yes BARENEED MA3 $ - Yes BARRD ISLANDS MA4 $ - Yes 10,000 BARTLETTS HARBOUR MA4 $ - Yes BAULINE MA3 $ - Yes BAY BULLS MA4 $ - Yes points across Canada BAY DE VERDE MA4 $ - Yes BAY L'ARGENT MA4 $ - Yes BAY ROBERTS MA3 $ - Yes BAYSIDE MA4 $ - Yes BAYTONA MA4 $ - Yes BAYVIEW MA4 $ - Yes BEACHES MA4 $ - Yes BEACHSIDE MA4 $ - Yes BEAR COVE MA4 $ - Yes BEAU BOIS MA3 $ - Yes BEAUMONT MA4 $ - Yes BELL ISLAND FRONT MA4 $ - Yes Notes: Some points not serviced daily. Shipments may be subject to a beyond charge if delivery is outside our regular service area. Standard Terms & Conditions will apply. Revised March 17, 2020 Newfoundland DIRECT SERVICE POINTS DANGEROUS CITY ZONE OFFLINE CHARGE GOODS SERVICE BELL ISLAND MA4 $ - Yes BELLBURNS MA4 $ - Yes BELLEORAM MA4 $ - Yes BELLEVUE MA4 $ - Yes shipping your envelopes, parcels and small skids to over BELLMANS COVE MA4 $ - Yes BENOITS COVE MA4 $ - Yes BENTON -

Codes Used in the Newfoundland Commercial and Recreational Fisheries
Environment Canada Environnement Canada •• Fisheries Service des peches and Marine Service et des sciences de la mer 1 DFO ll ll i ~ ~~ll[lflll ~i~ 1 \11 1f1i! l1[1li eque 07003336 Codes Used in the Newfoundland Commercial and Recreational Fisheries by Don E. Waldron Data Record Series No. NEW/D-74-2 Resource Development Branch Newtoundland Region ) CODES USED IN THE NEWFOUNDLAND COMMERCIAL AND RECREATIONAL FISHERIES by D.E. Waldron Resource Development Branch Newfoundland Region Fisheries & Marine Service Department of the Environment St. John's, N'fld. February, 1974 GULF FlSHERIES LIBRARY FISHERIES & OCEANS gwt.IV HEOUE DES PECHES GOLFE' PECHES ET OCEANS ABSTRACT Data Processing is used by most agencies involved in monitoring the recreational and commercial fisheries of Newfoundland. There are three Branches of the Department of the Environment directly involved in Data Collection and Processing. The first two are the Inspection and the Conservation and Protection Branches (the collectors) and the Economics and Intelligence Branch (the processors)-is the third. To facilitate computer processing, an alpha-numeric coding system has been developed. There are many varieties of codes in use; however, only species, gear, ICNAF area codes, Economic and Intelligence Branch codes, and stream codes will be dealt with. Figures and Appendices are supplied to help describe these codes. ii TABLE OF CONTENTS ABSTRACT ........... .. ... .... ... ........... ................ ii ACKNOWLEDGEMENTS iv LIST .or FIGURES ....... .................................... v LIST OF TABLES ............................................ vi INTRODUCTION l Description of Data Coding .............. ~ .. .... ... 3 {A) Coding Varieties ••••••••••••••• 3 (I) Species Codes 3 ( II ) Gear Codes 3 (III) Area Codes 3 (i) ICNKF 4 (ii) Statistical Codes 7 (a) Statistical Areas 7 (b) Statistical Sections 7 (c) Community (Settlement) Codes 17 (iii) Comparison of ICNAF AND D.O.E. -
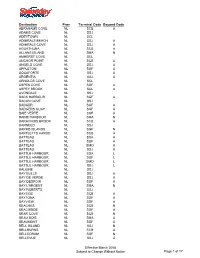
Destination Prov Terminal Code Beyond Code ABRAHAMS COVE
Destination Prov Terminal Code Beyond Code ABRAHAMS COVE NL SCB A ADAMS COVE NL SSJ ADEYTOWN NL SCL ADMIRALS BEACH NL SSJ A ADMIRALS COVE NL SSJ A AGUATHUNA NL SCB A ALLANS ISLAND NL SMA N AMHERST COVE NL SCL ANCHOR POINT NL SCB A ANGELS COVE NL SSJ A APPLETON NL SGF A AQUAFORTE NL SSJ A ARGENTIA NL SSJ A ARNOLDS COVE NL SCL ASPEN COVE NL SGF A ASPEY BROOK NL SCL A AVONDALE NL SSJ BACK HARBOUR NL SGF A BACON COVE NL SSJ BADGER NL SGF A BADGERS QUAY NL SGF A BAIE VERTE NL SGF A BAINE HARBOUR NL SMA N BARACHOIS BROOK NL SCB A BARNEED NL SSJ BARRD ISLANDS NL SGF N BARTLETTS HARBO NL SCB A BATTEAU NL SDA A BATTEAU NL SGF A BATTEAU NL SMO A BATTEAU NL SSJ A BATTLE HARBOUR NL SDA L BATTLE HARBOUR NL SGF L BATTLE HARBOUR NL SMO L BATTLE HARBOUR NL SSJ L BAULINE NL SSJ BAY BULLS NL SSJ A BAY DE VERDE NL SSJ A BAY DESPOIR NL SGF A BAY L'ARGENT NL SMA N BAY ROBERTS NL SSJ BAYSIDE NL SCB A BAYTONA NL SGF A BAYVIEW NL SGF A BEACHES NL SCB N BEACHSIDE NL SGF A BEAR COVE NL SCB N BEAU BOIS NL SMA A BEAUMONT NL SGF A BELL ISLAND NL SSJ N BELLBURNS NL SCB A BELLEORAM NL SGF N BELLEVUE NL SSJ A Effective March 2008 Subject to Change Without Notice Page 1 of 17 Destination Prov Terminal Code Beyond Code BELLMANS COVE NL SCB A BENOITS COVE NL SCB A BENTON NL SGF A BIDE ARM NL SCB N BIG BROOK NL SAN N BIRCHY BAY NL SGF A BIRCHY COVE BB NL SCL BIRCHY HEAD NL SCB N BIRD COVE NL SCB A BISCAY BAY NL SSJ BISHOPS COVE NL SSJ BISHOPS FALLS NL SGF BLACK DUCK NL SCB A BLACK DUCK COVE NL SCB A BLACK DUCK SIDI NL SCB A BLACK TICKLE NL SDA L BLACK TICKLE NL SGF L BLACK -

Newfoundland 3Ait Service a Cost Re Over Alternative
Summary Report NEWFOUNDLAND 3AIT SERVICE A COST RE OVER ALTERNATIVE SH 344.8 E. Gordon RodgErs B3 Atlantic Development Programs Branch R6 Draft 1 — Jury 31, 1980 209097 (SUMMARY REPORT NEWFOUNDLAND BAIT SERVICE A COST RECOVERY ALTERNATIVE E. Gordon Rodgers, A/Director Atlantic Development Programs Branch July 31, 1980 Draft 1 CONTENTS PAGE INTRODUCTION 1 GENERAL HISTORY 3 NEWFOUNDLAND BAIT SERVICE 5 Management and Organization 5 Facilities 6 Operations - Bait Supply and Uses 7 Bait Costs - Purchases and Sales 9 Maritimes - Bait Supply 11 Bait Facilities Operating Costs 11 NEED FOR THE NEWFOUNDLAND BAIT SERVICE 13 RECOMMENDATIONS FOR COST RECOVERY 14 CONCLUSIONS 16 APPENDICES Map 1 Distribution of Bait Depots 1949 18 Map 2 Distribution of Depots and Units 1979 19 Map 3 Distribution of Fish Landings and Values 1978 20 Map 4 Distribution of Crustacean Landings and Values 1978 21 Map 5 Distribution of Registered Fishermen 22 Table 1 Summary Historical Changes in Number of Facilities 23 Table 2 Depot History - List of Facilities (1949-1979) 24 Table 3 Unit History - List of Facilities (1949-1979) 25 Table 4 Indeterminate Positions 1979 26 Table 5 Person Year Allotments 27 Table 6 Depot Capacity and Age - 1979 28 Table 7 Unit Capacity and Age - 1979 29 Table 8 Depot - Opening and Closing Dates 1979-1980 30 Table 9 Unit - Opening and Closing Dates 1979-1980 31 Table 10 Depot Accessibility and Adjacent Plants 1979 32 Table 11 Unit Accessibility and Adjacent Plants 1979 33 Table 12 Landings and Values of Catches on -

Knights of Columbus Space Available for 3 New Weekly
RYAN’S ST. MARY’S TELEPHONE 525-2115 Before you purchase a new home, Grocery & Confectionary Weekly Specials Refinance or renew your mortgage. Bakery/Homemade Bread/ Cold Plates /Snack Bar Call me to discuss your options! Liquor Store/Gas bar Anne Marie Drohan Open: Monday-Saturday 8:00am – 9:00pm (709)764-4959 Sunday 10:00am – 8:00pm [email protected] NOW WITH FULL BACK UP POWER SUPPLY INCLUDING www.mortgageweb.ca/annmariedrohan OUR TAKE OUT TO SERVE YOU IN POWER OUTAGES Best Mortgage Rates Available! KNIGHTS OF COLUMBUS St. Mary’s Bay Council #13320 for information. Telephone 738-1234 Dave: 229-7395 Howard: 521-2431 Mike: 525-2551 Kevin: 521-2560 Two locations: 279 Portugal Cove Road, St. John’s 2 St. David’s Avenue, Mount Pearl www.oldtownpizzaria.com 155 McNamara Drive Paradise, NL A1L 0A7 www.noeldaleylaw.com Office Hours: 8:30 am-5:00 pm [email protected] Telephone: 781-3264 Fax: 781-3265 For a list of services: www.powervacnl.com Toll Free 1.855.375.6635 Newfoundland Office St. Joseph’s, Box 3 St. Mary’s Bay, NL, CA A0B 3A0 Res 709.521.2376 “The oldest and longest serving law office in St. Mary’s Bay” Royal Canadian Legion Branch 62 Father Val Power Centre Riverhead, S.M.B. Riverhead, Telephone # 709-525-2260 Telephone: 709-525-2712 Gym Hours: Mon – Thurs 9-11AM & 7-9PM Sun 7-9PM Business Hours: Monday – Friday 7:30 pm – 12:00 am Depot Hours: Tuesday & Thursday 12:30 – 2:30 PM Saturday 2pm-5pm & 7pm – 12 am Saturday 10:00am – 12 noon & 1:00 – 3:00pm Auction Monday night @ 8:30 pm sponsored by Ladies Auxiliary Friday Night – BINGO 8:30 pm – Jackpot / Loonie Jar / Lucky7 Mount Carmel Building Supplies Ltd.