Late Middle Eocene Primate from Myanmar and the Initial Anthropoid Colonization of Africa
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

For Early Anthropoids
EVOLUTIONARY ANTHROPOLOGY 24:41–42 (2015) NEWS Westward Ho! for Early Anthropoids etween March 17 and 19, The likely have been an ideal time for dis- quarries, discussed the chronological School for Advanced Research persals from Asia to Africa because of dates of the stratigraphic section, the Bin Santa Fe, New Mexico, the geographical positions of Middle adaptations of the many species, and hosted a short but intense, seminar/ Eastern tectonic blocks and the likeli- the ongoing debates about the phylo- workshop on “The Colonization of hood of high water volume flowing genetic relationships of different Africa by Early Anthropoid Primates.” southwest from large Asian rivers taxa. The seminar, organized by Chris toward Africa in conjunction with pre- Chris Beard (Kansas) reviewed the Beard (Kansas), involved two full and vailing westward currents. evidence of anthropoid evolution in intense days of presentations and dis- Gregoire Metais (Museum National Africa and Asia, then addressed the cussions, followed by a field trip to d’Histoire Naturelle, France) discussed importance of over-water dispersal Paleocene and Eocene localities in the “Potential Role of Turkey as a Bio- in primate evolution. He argued that northern New Mexico’s San Juan geographic Crossroads for Eurasian the phenomenon that is often called Basin. and African Mammals During the “sweepstakes dispersal” is not ran- Africa has been a major center for Paleogene.” Modern Turkey has an dom (and thus should perhaps be the evolution of anthropoid primates intricate geological history. Recent referred to as a special case of filter over at least the last 35 million years. paleontological work by Metais and dispersal). -

Late Middle Eocene Primate from Myanmar and the Initial Anthropoid Colonization of Africa
Late Middle Eocene primate from Myanmar and the initial anthropoid colonization of Africa Abstract Reconstructing the origin and early evolutionary history of anthropoid primates (monkeys, apes, and humans) is a current focus of paleoprimatology. Although earlier hypotheses frequently supported an African origin for anthropoids, recent discoveries of older and phylogenetically more basal fossils in China and Myanmar indicate that the group originated in Asia. Given the Oligocene-Recent history of African anthropoids, the colonization of Africa by early anthropoids hailing from Asia was a decisive event in primate evolution. However, the fossil record has so far failed to constrain the nature and timing of this pivotal event. Here we describe a fossil primate from the late middle Eocene Pondaung Formation of Myanmar, Afrasia djijidae gen. et sp. nov., that is remarkably similar to, yet dentally more primitive than, the roughly contemporaneous North African anthropoid Afrotarsius. Phylogenetic analysis suggests that Afrasia and Afrotarsius are sister taxa within a basal anthropoid clade designated as the infraorder Eosimiiformes. Current knowledge of eosimiiform relationships and their distribution through space and time suggests that members of this clade dispersed from Asia to Africa sometime during the middle Eocene, shortly before their first appearance in the African fossil record. Crown anthropoids and their nearest fossil relatives do not appear to be specially related to Afrotarsius, suggesting one or more additional episodes of dispersal from Asia to Africa. Hystricognathous rodents, anthracotheres, and possibly other Asian mammal groups seem to have colonized Africa at roughly the same time or shortly after anthropoids gained their first toehold there . -

8. Primate Evolution
8. Primate Evolution Jonathan M. G. Perry, Ph.D., The Johns Hopkins University School of Medicine Stephanie L. Canington, B.A., The Johns Hopkins University School of Medicine Learning Objectives • Understand the major trends in primate evolution from the origin of primates to the origin of our own species • Learn about primate adaptations and how they characterize major primate groups • Discuss the kinds of evidence that anthropologists use to find out how extinct primates are related to each other and to living primates • Recognize how the changing geography and climate of Earth have influenced where and when primates have thrived or gone extinct The first fifty million years of primate evolution was a series of adaptive radiations leading to the diversification of the earliest lemurs, monkeys, and apes. The primate story begins in the canopy and understory of conifer-dominated forests, with our small, furtive ancestors subsisting at night, beneath the notice of day-active dinosaurs. From the archaic plesiadapiforms (archaic primates) to the earliest groups of true primates (euprimates), the origin of our own order is characterized by the struggle for new food sources and microhabitats in the arboreal setting. Climate change forced major extinctions as the northern continents became increasingly dry, cold, and seasonal and as tropical rainforests gave way to deciduous forests, woodlands, and eventually grasslands. Lemurs, lorises, and tarsiers—once diverse groups containing many species—became rare, except for lemurs in Madagascar where there were no anthropoid competitors and perhaps few predators. Meanwhile, anthropoids (monkeys and apes) emerged in the Old World, then dispersed across parts of the northern hemisphere, Africa, and ultimately South America. -

Early Eocene Primates from Gujarat, India
ARTICLE IN PRESS Journal of Human Evolution xxx (2009) 1–39 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Early Eocene Primates from Gujarat, India Kenneth D. Rose a,*, Rajendra S. Rana b, Ashok Sahni c, Kishor Kumar d, Pieter Missiaen e, Lachham Singh b, Thierry Smith f a Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, USA b H.N.B. Garhwal University, Srinagar 246175, Uttarakhand, India c Panjab University, Chandigarh 160014, India d Wadia Institute of Himalayan Geology, Dehradun 248001, Uttarakhand, India e University of Ghent, B-9000 Ghent, Belgium f Royal Belgian Institute of Natural Sciences, B-1000 Brussels, Belgium article info abstract Article history: The oldest euprimates known from India come from the Early Eocene Cambay Formation at Vastan Mine Received 24 June 2008 in Gujarat. An Ypresian (early Cuisian) age of w53 Ma (based on foraminifera) indicates that these Accepted 8 January 2009 primates were roughly contemporary with, or perhaps predated, the India-Asia collision. Here we present new euprimate fossils from Vastan Mine, including teeth, jaws, and referred postcrania of the Keywords: adapoids Marcgodinotius indicus and Asiadapis cambayensis. They are placed in the new subfamily Eocene Asiadapinae (family Notharctidae), which is most similar to primitive European Cercamoniinae such as India Donrussellia and Protoadapis. Asiadapines were small primates in the size range of extant smaller Notharctidae Adapoidea bushbabies. Despite their generally very plesiomorphic morphology, asiadapines also share a few derived Omomyidae dental traits with sivaladapids, suggesting a possible relationship to these endemic Asian adapoids. In Eosimiidae addition to the adapoids, a new species of the omomyid Vastanomys is described. -

Evidence for an Asian Origin of Stem Anthropoids
Evidence for an Asian origin of stem anthropoids Richard F. Kay1 Department of Evolutionary Anthropology, Duke University, Durham, NC 27708-03083 n PNAS, Chaimanee et al. (1) report Genetic, embryological, and anatomical We are not there yet. Recent comprehen- a previously undescribed species of evidence demonstrates that the sister sive phylogenetic analyses stemming from I primate, Afrasia, from the late Middle group of Anthropoidea is the south Asian virtually the same datasets yield somewhat Eocene of Burma. They identify tarsier, with the two forming the crown different cladograms that reflect sensitivity Afrasia as the sister taxon to the African group Haplorhini (6–8). The other clade of to which taxa are included in the analysis genus Afrotarsius but slightly more primitive extant primates is the lemurs and lorises, and which sets of analytical assumptions are than it and allied with stem Anthropoidea called Strepsirrhini. Eocene Holarctic selected. Pertinent to the biogeographic of south Asia. Anthropoidea is the taxo- Omomyoidea are generally considered as conclusions of Chaimanee et al. (1), Seiffert nomic group that today includes New and stem haplorhines, although the precise et al. (11) conclude that Afrotarsius cha- Old World monkeys, apes, and humans. If relationship of omomyoids to tarsiers and trathi [a younger species than the one that upheld, the biogeographic significance of anthropoids is uncertain (8). Tarsius often Chaimanee et al. (1) describe] is an African these results is profound: If Afrasia and is considered to be a relictual omomyoid tarsioid. If Seiffert et al. (11) are correct, Afrotarsius are as closely related as Chai- in south Asia. -

Evolutionary History of Lorisiform Primates
Evolution: Reviewed Article Folia Primatol 1998;69(suppl 1):250–285 oooooooooooooooooooooooooooooooo Evolutionary History of Lorisiform Primates D. Tab Rasmussen, Kimberley A. Nekaris Department of Anthropology, Washington University, St. Louis, Mo., USA Key Words Lorisidae · Strepsirhini · Plesiopithecus · Mioeuoticus · Progalago · Galago · Vertebrate paleontology · Phylogeny · Primate adaptation Abstract We integrate information from the fossil record, morphology, behavior and mo- lecular studies to provide a current overview of lorisoid evolution. Several Eocene prosimians of the northern continents, including both omomyids and adapoids, have been suggested as possible lorisoid ancestors, but these cannot be substantiated as true strepsirhines. A small-bodied primate, Anchomomys, of the middle Eocene of Europe may be the best candidate among putative adapoids for status as a true strepsirhine. Recent finds of Eocene primates in Africa have revealed new prosimian taxa that are also viable contenders for strepsirhine status. Plesiopithecus teras is a Nycticebus- sized, nocturnal prosimian from the late Eocene, Fayum, Egypt, that shares cranial specializations with lorisoids, but it also retains primitive features (e.g. four premo- lars) and has unique specializations of the anterior teeth excluding it from direct lorisi- form ancestry. Another unnamed Fayum primate resembles modern cheirogaleids in dental structure and body size. Two genera from Oman, Omanodon and Shizarodon, also reveal a mix of similarities to both cheirogaleids and anchomomyin adapoids. Resolving the phylogenetic position of these Africa primates of the early Tertiary will surely require more and better fossils. By the early to middle Miocene, lorisoids were well established in East Africa, and the debate about whether these represent lorisines or galagines is reviewed. -

Effects of Additional Postcranial Data on the Topologies of Early Eocene Primate
Effects of Additional Postcranial Data on the Topologies of Early Eocene Primate Phylogenies Julia Stone ANT 679HB Special Honors in the Department of Anthropology The University of Texas at Austin May 2021 Liza J. Shapiro Department of Anthropology Supervising Professor E. Christopher Kirk Department of Anthropology Second Reader Acknowledgements This project would not have been completed without the help of my thesis supervisor Dr. Liza Shapiro. I was inspired to start the project after taking her courses in Primate Evolution and Primate Anatomy. She also met with me many times throughout this year to discuss the trajectory of the project as well as discuss papers that became the foundation of the thesis. She assisted me through technology issues, unresolved trees, and many drafts, and she provided invaluable guidance throughout this process. I also want to thank Dr. Christopher Kirk for being my second reader and providing extremely helpful insight into how to write about phylogenetic results. He also helped me to be more specific and accurate in all of the sections of the thesis. I learned a lot about writing theses in general due to his comments on my drafts. I also truly appreciate the help of Ben Rodwell, a graduate student at the University of Texas at Austin. He provided excellent help and support regarding the Mesquite and TNT software used in this project. ii Effects of Additional Postcranial Data on the Topologies of Early Eocene Primate Phylogenies by Julia Stone, BA The University of Texas at Austin SUPERVISOR: Liza J. Shapiro There are multiple hypotheses regarding the locomotor behaviors of the last common ancestor to primates and which fossil primates best represent that ancestor. -

1 Old World Monkeys
2003. 5. 23 Dr. Toshio MOURI Old World monkey Although Old World monkey, as a word, corresponds to New World monkey, its taxonomic rank is much lower than that of the New World Monkey. Therefore, it is speculated that the last common ancestor of Old World monkeys is newer compared to that of New World monkeys. While New World monkey is the vernacular name for infraorder Platyrrhini, Old World Monkey is the vernacular name for superfamily Cercopithecoidea (family Cercopithecidae is limited to living species). As a side note, the taxon including Old World Monkey at the same taxonomic level as New World Monkey is infraorder Catarrhini. Catarrhini includes Hominoidea (humans and apes), as well as Cercopithecoidea. Cercopithecoidea comprises the families Victoriapithecidae and Cercopithecidae. Victoriapithecidae is fossil primates from the early to middle Miocene (15-20 Ma; Ma = megannum = 1 million years ago), with known genera Prohylobates and Victoriapithecus. The characteristic that defines the Old World Monkey (as synapomorphy – a derived character shared by two or more groups – defines a monophyletic taxon), is the bilophodonty of the molars, but the development of biphilophodonty in Victoriapithecidae is still imperfect, and crista obliqua is observed in many maxillary molars (as well as primary molars). (Benefit, 1999; Fleagle, 1999) Recently, there is an opinion that Prohylobates should be combined with Victoriapithecus. Living Old World Monkeys are all classified in the family Cercopithecidae. Cercopithecidae comprises the subfamilies Cercopithecinae and Colobinae. Cercopithecinae has a buccal pouch, and Colobinae has a complex, or sacculated, stomach. It is thought that the buccal pouch is an adaptation for quickly putting rare food like fruit into the mouth, and the complex stomach is an adaptation for eating leaves. -
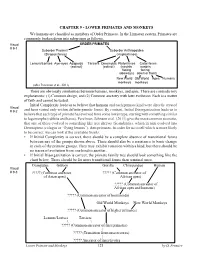
CHAPTER 9 - LOWER PRIMATES and MONKEYS We Humans Are Classified As Members of Order Primates
CHAPTER 9 - LOWER PRIMATES AND MONKEYS We humans are classified as members of Order Primates. In the Linnaean system, Primates are commonly broken down into subgroups as follows. Visual ORDER PRIMATES # 9-1 Suborder Prosimii Suborder Anthropoidea (Strepsirrhines) (Haplorrhines) Lemurs/Lorises Aye-ayes Adapoids Tarsiers Omomyids Platyrrhines Catarrhines (extinct) (extinct) (nostrils nostrils facing facing sideways) down or front) New World Old World Apes Humans monkeys monkeys (after Perelman et al., 2011) There are obviously similarities between humans, monkeys, and apes. There are contradictory explanations: (1) Common design, and (2) Common ancestry with later evolution. Each is a matter of faith and cannot be tested. Initial Complexity leads us to believe that humans and each primate kind were directly created Visual # 9-2 and have varied only within definite genetic limits. By contrast, Initial Disorganization leads us to believe that each type of primate has evolved from some lower type, starting with something similar to lagomorphs (rabbits and hares). Perelman, Johnson et al. (2011) give the most common scenario, that one of these evolved to something like tree shrews (Scandentia), which in turn evolved into Dermoptera (colugos or “flying lemurs”), then primates. In order for us to tell which is more likely to be correct, we can look at the available fossils. • If Initial Complexity is correct, there should be a complete absence of transitional forms between any of the groups shown above. There should also be a resistance to basic change in each of the primate groups. They may exhibit variation within a kind, but there should be no traces of evolution from one kind to another. -

Science Journals — AAAS
RESEARCH ◥ differ from those of Wailekia and Kyitchaungia RESEARCH ARTICLES in having lingually open trigonids with proto- conid and metaconid more closely approximated and talonids bearing more trenchant cristids. PRIMATE EVOLUTION UppermolarsdifferfromthoseofGuangxilemur in having more prominent parastyles and pre- and postprotocristae joining the bases of the Oligocene primates from China reveal paracone and metacone, respectively, rather than merging with the pre- and postcingula. Upper molars with weaker buccal cingulum and incom- divergence between African and plete mesostyle, in contrast to Guangxilemur tongi. Lower molars differ from those of Guang- Asian primate evolution xilemur singsilai in having lingually open trig- onids and deeper, more extensive valley separating Xijun Ni,1,2* Qiang Li,1,2 Lüzhou Li,1 K. Christopher Beard3,4 entoconid and hypoconulid. Differs from Miocene 4 sivaladapids in lacking fully molariform P4 and P Profound environmental and faunal changes are associated with climatic deterioration and having upper molars with distinct pericone during the Eocene-Oligocene transition (EOT) roughly 34 million years ago. Reconstructing and hypocone cusps and pre- and postprotocris- how Asian primates responded to the EOT has been hindered by a sparse record of tae joining bases of paracone and metacone. Oligocene primates on that continent. Here, we report the discovery of a diverse primate Etymology: Generic name recognizes the geo- fauna from the early Oligocene of southern China. In marked contrast to Afro-Arabian graphic provenance of this taxon and its adapi- Downloaded from Oligocene primate faunas, this Asian fauna is dominated by strepsirhines. There appears to form affinities. be a strong break between Paleogene and Neogene Asian anthropoid assemblages. -

Rapid Laurasian Diversification of a Pantropical Bird Family During The
Ibis (2020), 162, 137–152 doi: 10.1111/ibi.12707 Rapid Laurasian diversification of a pantropical bird family during the Oligocene–Miocene transition CARL H. OLIVEROS,1,2* MICHAEL J. ANDERSEN,3 PETER A. HOSNER,4,7 WILLIAM M. MAUCK III,5,6 FREDERICK H. SHELDON,2 JOEL CRACRAFT5 & ROBERT G. MOYLE1 1Department of Ecology and Evolutionary Biology and Biodiversity Institute, University of Kansas, Lawrence, KS, USA 2Museum of Natural Science and Department of Biological Sciences, Louisiana State University, Baton Rouge, LA, USA 3Department of Biology and Museum of Southwestern Biology, University of New Mexico, Albuquerque, NM, USA 4Division of Birds, Department of Vertebrate Zoology, National Museum of Natural History, Washington, DC, USA 5Department of Ornithology, American Museum of Natural History, New York, NY, USA 6New York Genome Center, New York, NY, USA 7Natural History Museum of Denmark and Center for Macroecology, Evolution, and Climate; University of Copenhagen, Copenhagen, Denmark Disjunct, pantropical distributions are a common pattern among avian lineages, but dis- entangling multiple scenarios that can produce them requires accurate estimates of his- torical relationships and timescales. Here, we clarify the biogeographical history of the pantropical avian family of trogons (Trogonidae) by re-examining their phylogenetic rela- tionships and divergence times with genome-scale data. We estimated trogon phylogeny by analysing thousands of ultraconserved element (UCE) loci from all extant trogon gen- era with concatenation and coalescent approaches. We then estimated a time frame for trogon diversification using MCMCTree and fossil calibrations, after which we performed ancestral area estimation using BioGeoBEARS. We recovered the first well-resolved hypothesis of relationships among trogon genera. -

The Tempo of Trophic Evolution in Small-Bodied Primates 2 3 4 Running Title: Trophic Evolution in Small-Bodied Primates 5 6 7 Jeremiah E
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.17.996207; this version posted March 20, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. 1 1 The tempo of trophic evolution in small-bodied primates 2 3 4 Running title: Trophic evolution in small-bodied primates 5 6 7 Jeremiah E. Scott 8 Department of Medical Anatomical Sciences 9 College of Osteopathic Medicine of the Pacific 10 Western University of Health Sciences 11 309 East Second Street 12 Pomona, California 91766 13 U.S.A. 14 [email protected] 15 16 17 Abstract 18 19 Objectives: As a primary trophic strategy, insectivory is uncommon and unevenly 20 distributed across extant primates. This pattern is partly a function of the challenges that 21 insectivory poses for large-bodied primates. In this study, I demonstrate that the uneven 22 distribution is also a consequence of variation in the rate of trophic evolution among small- 23 bodied lineages. 24 Methods: The sample consisted of 307 species classified by primary trophic strategy and 25 body size, creating an ordered three-state character: small-insectivorous, small-herbivorous, 26 and large-herbivorous. I tested for rate heterogeneity by partitioning major clades from the 27 rest of the primate tree and estimating separate rates of transition between herbivory and 28 insectivory for small-bodied lineages in each partition. 29 Results: Bayesian analysis of rate estimates indicates that a model with two rates of trophic 30 evolution provides the best fit to the data.