Asellidae) from Southwestern Virginia, with Notes on Its Ecology and Additional Cave Records for the Genus in the Appalachians
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Natur Und Heimat
Natur u. Heimat, 37. Jahrg., Heft 3, 1977 (1968): Gehäuse von Insekten-Larven, insbesondere von Chironomiden, in quar tären Sedimenten. Mitt. Geol. Inst. Univers. Hannover, 8, 34-53. ___.:._ HrLTERMANN, H. (1975): Kleiner Führer durch Solbad Laer T. W. Suderberger Hefte 1. - HrL TERMANN, H . (1976): Ein vergessener mittelalterlicher Baustein. Jb. Heimatbund Osnabrück-Land, 54-59. - HrLTERMANN, H. & K. MÄDLER (1977): Charophyten als palökologische Indikatoren und ihr Vorkommen in den Sinterkalken von Bad Laer T. W. Paläontol. Z. (im Druck). - ZEISSLER, H. (1977): Konchylien aus dem holozänen Travertin von Bad Laer, Kreis Osnabrück. (im Druck). Anschrift des Verfassers1: Prof. Dr. H. Hiltermann, Milanring 11, D-4518 Bad Laer. Die ersten Nachweise der Wasserassel Proasellus meridianus CRacovitza, 1919) (Crustacea, Isopoda Asellidae) im Einzugsgebiet der Ems KARL FRIEDRICH HERHAUS, Münster In Deutschland ist die von HENRY und MAGNIEZ (1970) revidierte Familie Asellida,e Sars, 1899, mit drei oberirdischen Arten vertreten. Die am weitesten verbreitete Art ist Asellus ( Asellus) aquaticus (L., 175 8); sie ist ein sibirisches Faunenelement, das sich postglazial nach Westen hin ausgebreitet hat (BrRSTEIN 1951; WILLIAMS 1962). Weitaus weniger häufig tritt die zweite Art, Proasellus coxalis (Dollfus, 1892), auf; diese im übrigen circummediterran verbreitete Art ist in Mittel europa mit der Unterart septentrionalis (Herbst, 1956) vertreten, die vermutlich erst in jüngster Zeit eingeschleppt worden ist (HERHAUS 1977). Am seltensten ist auf deutschem Boden die dritte Art, Proasellus meridianus (Racovitza, 1919). P. meridianus ist eine autochthon west europä,isch-atlantische Form (GRUNER 1965); in Deutschland wurde sie von STAMMER (1932) am linken Niederrhein nachgewiesen. Für die sichere Bestimmung der drei Arten ist die Untersuchung der Pleopoden II unter dem Binokular unerläßlich. -

Species of Greatest Conservation Need
APPENDIX A. VIRGINIA SPECIES OF GREATEST CONSERVATION NEED Taxa Common Scientific Name Tier Cons. Opp. Habitat Descriptive Habitat Notes Name Ranking Amphibians Barking Hyla gratiosa II a Forest Forests near or within The Virginia Fish and Wildlife Information System indicates treefrog shallow wetlands the loss suitable wetlands constitute the greatest threats to this species. DGIF recommends working to maintain or restore forested buffers surrounding occupied wetlands. These needs are consistent with action plan priorities to conserve and restore wetland habitats and associated buffers. Recently discovered populations within its known range, may indicate this species is more abundant than previously believed. An in-depth investigation into its status may warrant delisting. This species will be prioritized as Tier 2a. Amphibians Blue Ridge Desmognathus IV c Forest High elevation seeps, This species' distribution is very limited. Other than limiting dusky orestes streams, wet rock faces, logging activity in the occupied areas, no conservation salamander and riparian forests actions have been identified. Unless other threats or actions are identified, this species will be listed as Tier 4c. Amphibians Blue Ridge Eurycea III a Wetland Mountain streams and The needs of this species are consistent with priorities for two-lined wilderae adjacent riparian areas maintaining and enhancing riparian forests and aquatic salamander with mixed hardwood or habitats. This species will be listed as Tier 3a. spruce-fir forests up to 6000 feet. Amphibians Carpenter Lithobates III a Wetland Freshwater wetlands with The needs of this species are consistent with action plan frog virgatipes sphagnum moss priorities to preserve and restore aquatic and wetland habitats and water quality. -

1994 IUCN Red List of Threatened Animals
The lUCN Species Survival Commission 1994 lUCN Red List of Threatened Animals Compiled by the World Conservation Monitoring Centre PADU - MGs COPY DO NOT REMOVE lUCN The World Conservation Union lo-^2^ 1994 lUCN Red List of Threatened Animals lUCN WORLD CONSERVATION Tile World Conservation Union species susvival commission monitoring centre WWF i Suftanate of Oman 1NYZ5 TTieWlLDUFE CONSERVATION SOCIET'' PEOPLE'S TRISr BirdLife 9h: KX ENIUNGMEDSPEaES INTERNATIONAL fdreningen Chicago Zoulog k.J SnuicTy lUCN - The World Conservation Union lUCN - The World Conservation Union brings together States, government agencies and a diverse range of non-governmental organisations in a unique world partnership: some 770 members in all, spread across 123 countries. - As a union, I UCN exists to serve its members to represent their views on the world stage and to provide them with the concepts, strategies and technical support they need to achieve their goals. Through its six Commissions, lUCN draws together over 5000 expert volunteers in project teams and action groups. A central secretariat coordinates the lUCN Programme and leads initiatives on the conservation and sustainable use of the world's biological diversity and the management of habitats and natural resources, as well as providing a range of services. The Union has helped many countries to prepare National Conservation Strategies, and demonstrates the application of its knowledge through the field projects it supervises. Operations are increasingly decentralised and are carried forward by an expanding network of regional and country offices, located principally in developing countries. I UCN - The World Conservation Union seeks above all to work with its members to achieve development that is sustainable and that provides a lasting Improvement in the quality of life for people all over the world. -

Asellota, Asellidae)
The Freshwater lsopods of the Genus Lirceus (Asellota, Asellidae) LESLIE HUBRICHT and J. G. MACKIN V Reprinted from "THE AMERICAN MIDLAND NATURALIST" Vol. 42, No. 2, pp. 334-349, September, 1949 Notre Dame, Ind. The Freshwater lsopods of the Genus Lirceus (Asellota, Asellidae) Leslie Hubricht 912 Main St., Danville, Virginia J. G. Mackin Texas A & M Research Foundation, Grande Isle, Louisiana In the genus Asellus we have, in the first and second pleopoda of the male, characters which can be relied upon to distinguish the species, but in Lirceus these appendages are so similar in different species that they are, with one exception, useless as a means of separation. This is not because the species of Lirceus are less distinct than Asellus, but requires that the animal as a whole be studied rather than a single appendage. To an experienced observer the various species are easily distinguished in the field by those subtile differences in form and color which are almost impossible to describe. Difficulties begin when one brings specimens into the laboratory and attempts to define the dif- ferences. The commonest cause of difficulty is depauperate specimens. Under adverse conditions the animals are stunted, the gnathopoda do not develop their processes, and the uropoda remain in juvenile form. When this depau- perization is extreme the collection cannot be determined, but must be as- sumed to belong to the species in the range of which it was found. The isopods discussed herein have, in the past, been treated under the generic name Mancasellus Harger, rather than the prior Lirceus of Rafinesque. -

Isopoda, Asellota) with Some Phylogenetic Considerations Concerning Asellota
Crustaceana 48 (2) 1985, E. J. Brill, Leiden A CONTRIBUTION TO THE GENERAL MORPHOLOGY OF PROTOJANIRIDAE (ISOPODA, ASELLOTA) WITH SOME PHYLOGENETIC CONSIDERATIONS CONCERNING ASELLOTA BY B. SKET Institut za biologijo, Univerza E. Kardelja v Ljubljani, p.p. 141, YU 61001 Ljubljana, Yugoslavia Since most representatives of the superfamily Gnathostenetroidoidea have been described only for "microtaxonomical" purposes, some important features and even peculiarities of the species and probably of the entire group have been overlooked. I took advantage of possessing a good sample of the comparatively very large representative, viz., Enckella lucei major Sket, 1982 (as well as some other species of the family Protojaniridae) to demonstrate some such features. Unfortunately, of the family Gnathostenetroididae I could study only specimens of an undescribed species from the Bahamas, while comparison with other representatives was possible only through illustrations. I abstained from comparing Enckella with the numerous and diverse Janiroidea, leaving this to a more versed student of that special group. I also abstained from discussing the remarkable idea about possible Asellotan nature of Microcerberidae (WageJe, J 980) which is anyway of little importance for the present subject. Cephalothorax ("head"). — With the exception of having two pairs of antennae, the Enckella's head bears greater resemblance to a prognathous in sect head than to an Asellid. The cephalic capsula is closed to a great extent, incorporating completely the first thoracomere. The capsula is closed and slightly sclerotised also behind the insertions of the maxillipeds, uniting them with the set of other mouth appendages. The clypeolabrum protrudes for wards, its inclination downwards is much less expressed than in most other groups of Isopoda. -

•Not! Ptdition, X:213-230
78 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY Carman, S. Knjyer, H. 1889. Cave Animal j from South we item Missouri. Bulle 1838. Gronlands Amphipoder. Kongelige danske viden* tin of the Museum of Comparative Zoology, skabernes Selskabbiologiske Skrifter, 7:229-326. 17(6):225-239. Latreille, P. A. Geoffrey Saint-Hilaire, E. L. 1803. Histoire naturelle, ginirale et particuliire, des 1764. Histoire abrigie dts Insectes qui se troitvcnt aux Cruslacis et dts Insectes, 6:1-391. Paris.* environs de Paris. Pari*.* Leonard, A. B,, and L. H. Ponder Gould, A. A, 1949. Crustacea of Eastern Kansas. Transactions of the 1841. Report on the Invertebrate Animals of Massachu Kansas Academy of Science, 52:168-204. setts. Cambridge: Massachusetts. Levi, H. W. Gruner, H. £. 1949. Two New Species of Cave Isopods from Pennsyl 1965. Die Tierwelt Deutschlands. Krebstiere oder Crus vania. Notulae Naturae, 220:1-fi. tacea. V, Isopoda. 53. Teil, 2 Lf. Jena. Linnaeus, C. Hansen, H. J. 1758. Systema Naturae. 10th Edition, 824 pages* 1888. Oversigt over det vestlige Gronlands Fauna. Mac Lin, J. G. Videnskabelige Meddeletier jra dansk Naturhis- 1940. A Key to the Oklahoma Species of the Family torisk Forening Kjobenhacn, 1887:177-198. Asellidae. Proceedings of the Oklahoma Academy Harford, W. G. W. of Science, 20:17-18. 1877. Description of a New Genus and Three New Spe Mackin, J. G, and L. Hubricht cies of Sessile Eyed Crustacea. Proceedings of the 1938. Records of Distribution of Species of Isopods in California Academy of Science, 7:53-55. Central and Southern United States, with Descrip Harger, O. tions of Four New Species of Mancasellus and 1874. -

Identification of Freshwater Invertebrates
Identification of Freshwater Invertebrates © 2008 Pennsylvania Sea Grant To request copies, please contact: Sara Grisé email: [email protected] Table of Contents A. Benthic Macroinvertebrates……………………….………………...........…………1 Arachnida………………………………..………………….............….…2 Bivalvia……………………...…………………….………….........…..…3 Clitellata……………………..………………….………………........…...5 Gastropoda………………………………………………………..............6 Hydrozoa………………………………………………….…………....…8 Insecta……………………..…………………….…………......…..……..9 Malacostraca………………………………………………....…….…....22 Turbellaria…………………………………………….….…..........…… 24 B. Plankton…………………………………………...……….………………............25 Phytoplankton Bacillariophyta……………………..……………………...……….........26 Chlorophyta………………………………………….....…………..........28 Cyanobacteria…...……………………………………………..…….…..32 Gamophyta…………………………………….…………...….…..…….35 Pyrrophycophyta………………………………………………………...36 Zooplankton Arthropoda……………………………………………………………....37 Ciliophora……………………………………………………………......41 Rotifera………………………………………………………………......43 References………………………………………………………….……………….....46 Taxonomy is the science of classifying and naming organisms according to their characteris- tics. All living organisms are classified into seven levels: Kingdom, Phylum, Class, Order, Family, Genus, and Species. This book classifies Benthic Macroinvertebrates by using their Class, Family, Genus, and Species. The Classes are the categories at the top of the page in colored text corresponding to the color of the page. The Family is listed below the common name, and the Genus and Spe- cies names -

And Community Structure
studet reasol food , and ir comp fauna and S the re and I classr other unde paras Th simp SpeciesInteractions this r inter and CommunityStructure short from Ti Questions concerning the nature and importance of species interac- vari< tions have become increasingly controversial. In the mid-1960sit was tive thought that interspecific competition was the major organizing factor ond in many communities. Through the pioneering work of the late Robert and MacArthur and his students,many aspectsof the structure and dynam- com ics of bird communities seemedbest explained by competitive interac- ied r tions, which could be describedin a generalway by the Lotka-Volterra spec competition equations. This work, or at least its generality, has been ofn challengedon three levels. First, there is the question of how important species interactions are in general and whether the interactions are competitive, parasitic, predatory, or mutualistic. For example, Connell whi (1975)noted that a significantfraction of, but by no means all, commu- nities are physically rather than biotically controlled. In addition, there As has been a tendency in the last few years to concentrate on single- dyn species demography. While interactions between species can still be the considered in the guise of age-specificfecundity and mortality effects, (va the emphasisis away from a coevolutionary perspective and toward a hos individualistic concept of communities. the Second, the question of which type of interaction is most important sor in structuring a particular community is very much open. Many ecol- tha ogists have pointed out that the birds studied by MacArthur and his wit SPECIESINTERACTIONS 97 studentswere at or near the top of the food chain. -

Endemic Flora and Fauna of Arkansas Henry W
Journal of the Arkansas Academy of Science Volume 36 Article 17 1982 Endemic Flora and Fauna of Arkansas Henry W. Robison Southern Arkansas University Kenneth L. Smith Arkansas Natural Heritage Commission Follow this and additional works at: http://scholarworks.uark.edu/jaas Part of the Terrestrial and Aquatic Ecology Commons Recommended Citation Robison, Henry W. and Smith, Kenneth L. (1982) "Endemic Flora and Fauna of Arkansas," Journal of the Arkansas Academy of Science: Vol. 36 , Article 17. Available at: http://scholarworks.uark.edu/jaas/vol36/iss1/17 This article is available for use under the Creative Commons license: Attribution-NoDerivatives 4.0 International (CC BY-ND 4.0). Users are able to read, download, copy, print, distribute, search, link to the full texts of these articles, or use them for any other lawful purpose, without asking prior permission from the publisher or the author. This Article is brought to you for free and open access by ScholarWorks@UARK. It has been accepted for inclusion in Journal of the Arkansas Academy of Science by an authorized editor of ScholarWorks@UARK. For more information, please contact [email protected], [email protected]. Journal of the Arkansas Academy of Science, Vol. 36 [1982], Art. 17 THE ENDEMIC FLORA AND FAUNA OF ARKANSAS HENRY W. ROBISON Department of Biological Sciences Southern Arkansas University Magnolia, Arkansas 71753 KENNETH L.SMITH Arkansas Natural Heritage Commission Suite 500, Continental Building Little Rock, Arkansas 72201 ABSTRACT Arkansas has an amazing diversity of plants and animals contained within its political boundaries. Forty-seven taxa are reported as Arkansas endemics, including seven plants, thirteen crustaceans (two amphipods, three isopods, eight crayfishes), nine insects (one mayfly, one caddisfly, three stoneflies, four beetles), ten snails, six fishes, and two salamanders. -

Native and Exotic Amphipoda and Other Peracarida in the River Meuse: New Assemblages Emerge from a Fast Changing Fauna
Hydrobiologia (2005) 542:203–220 Ó Springer 2005 H. Segers & K. Martens (eds), Aquatic Biodiversity II DOI 10.1007/s10750-004-8930-9 Native and exotic Amphipoda and other Peracarida in the River Meuse: new assemblages emerge from a fast changing fauna Guy Josens1,*, Abraham Bij de Vaate2, Philippe Usseglio-Polatera3, Roger Cammaerts1,4, Fre´de´ric Che´rot1,4, Fre´de´ric Grisez1,4, Pierre Verboonen1 & Jean-Pierre Vanden Bossche1,4 1Universite´ Libre de Bruxelles, Service de syste´matique et d’e´cologie animales, av. Roosevelt, 50, cp 160/13, B-1050 Bruxelles 2Institute for Inland Water Management & Waste Water Treatment, Lelystad, The Netherlands 3Universite´ de Metz, LBFE, e´quipe de De´moe´cologie, av. Ge´ne´ral Delestraint, F-57070 Metz 4Centre de Recherche de la Nature, des Foreˆts et du Bois, DGRNE, Ministe`re de la Re´gion wallonne, Avenue Mare´chal Juin, 23, B-5030 Gembloux (*Author for correspondence: e-mail: [email protected]) Key words: aquatic biodiversity, alien species, invasive species, invasibility, community dynamics, Dikerogammarus villosus, Chelicorophium curvispinum Abstract Samples issued from intensive sampling in the Netherlands (1992–2001) and from extensive sampling carried out in the context of international campaigns (1998, 2000 and 2001) were revisited. Additional samples from artificial substrates (1992–2003) and other techniques (various periods) were analysed. The combined data provide a global and dynamic view on the Peracarida community of the River Meuse, with the focus on the Amphipoda. Among the recent exotic species found, Crangonyx pseudogracilis is regressing, Dikerogammarus haemobaphes is restricted to the Condroz course of the river, Gammarus tigrinus is restricted to the lowlands and seems to regress, Jaera istri is restricted to the ‘tidal’ Meuse, Chelicorophium curvispinum is still migrating upstream into the Lorraine course without any strong impact on the other amphipod species. -
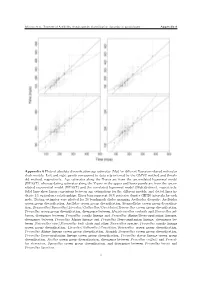
Appendix 6 Plots of Absolute Diversification Age Estimates (Ma
Morvan et al. Timetree of Aselloidea reveals species diversification dynamics in groundwater Appendix 6 Appendix 6 Plots of absolute diversification age estimates (Ma) for different Bayesian relaxed molecular clock models. Left and right panels correspond to data sets inferred by the GMYC method and thresh- old method, respectively. Age estimates along the X-axis are from the uncorrelated lognormal model (BEAST), whereas dating estimates along the Y-axis in the upper and lower panels are from the uncor- related exponential model (BEAST) and the correlated lognormal model (Multidivtime), respectively. Solid lines show linear regressions between age estimations for the different models, and dotted lines in- dicate 1:1 equivalence relationships. Error bars represent 95% posterior density (HPD) intervals for each node. Dating estimates were plotted for 20 benchmark clades spanning Aselloidea diversity: Aselloidea crown group diversification, Asellidae crown group diversification, Stenasellidae crown group diversifica- tion, Bragasellus/Synasellus/Lirceolus/Gallasellus/Caecidotea/Remasellus crown group diversification, Proasellus crown group diversification, divergence between Mexistenasellus coahuila and Stenasellus gal- hanoe, divergence between Proasellus coxalis lineage and Proasellus Alpine/Ibero-aquitanian lineages, divergence between Proasellus Alpine lineage and Proasellus Ibero-aquitanian lineage, divergence be- tween Stenasellus virei/Stenasellus buili clade and other Stenasellus species, Proasellus coxalis lineage crown group diversification, Lirceolus/Gallasellus/Caecidotea/Remasellus crown group diversification, Proasellus Alpine lineage crown group diversification, Spanish Bragasellus crown group diversification, Proasellus Ibero-aquitanian lineage crown group diversification, Proasellus slavus lineage crown group diversification, Asellus crown group diversification, divergence between Proasellus coiffaiti and Proasel- lus slovenicus, Synasellus crown group diversification, and divergence between Proasellus beroni and Proasellus ligusticus. -

II. Sampling Design
National Park Service U.S. Department of the Interior Natural Resource Program Center Protocol for Monitoring Aquatic Invertebrates of Small Streams in the Heartland Inventory & Monitoring Network Natural Resource Report NPS/HTLN/NRR—2008/042 A Heartland Network Monitoring Protocol protecting the habitat of our heritage i ON THE COVER Herbert Hoover birthplace cottage at Herbert Hoover NHS, prescribed fire at Tallgrass Prairie NPres, aquatic invertebrate monitoring at George Washington Carver NM, the Mississippi River at Effigy Mounds NM. ii Protocol for Monitoring Aquatic Invertebrates of Small Streams in the Heartland Inventory & Monitoring Network David E. Bowles, Michael H. Williams, Hope R. Dodd, Lloyd W. Morrison, Janice A. Hinsey, Catherine E. Ciak, Gareth A. Rowell, Michael D. DeBacker, and Jennifer L. Haack National Park Service Heartland I&M Network Wilson’s Creek National Battlefield 6424 West Farm Road 182 Republic, Missouri 65738 June 2008 U.S. Department of the Interior National Park Service Natural Resource Program Center Fort Collins, Colorado i The Natural Resource Publication series addresses natural resource topics that are of interest and applicability to a broad readership in the National Park Service and to others in the management of natural resources, including the scientific community, the public, and the NPS conservation and environmental constituencies. Manuscripts are peer- reviewed to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and is designed and published in a professional manner. Natural Resource Reports are the designated medium for disseminating high priority, current natural resource management information with managerial application. The series targets a general, diverse audience, and may contain NPS policy considerations or address sensitive issues of management applicability.