Seedless Mechanism of a New Citrus Cultivar 'Huami Wuhegonggan' (Citrus Sinensis × C. Reticulata)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Social Assessment Report
IPP574 v2 World Bank Loan Public Disclosure Authorized Hunan Forest Restoration and Development Project (HFRDP) Social Assessment Report Public Disclosure Authorized Public Disclosure Authorized Hunan Provincial Forest Foreign Fund Project Management Office Public Disclosure Authorized Social Assessment Team of HFRDP March, 2012 Social Assessment Report for Hunan Forest Restoration and Development Project Abbreviations CFB: County Forestry Bureau FC: Forest Cooperative HFRDP: Hunan Forest Restoration and Development Project HH: household HN: Hunan Province PCP: Participatory consultation and planning PFD: Provincial Forestry Department PPMO: Provincial Project Management Office PRA: Participatory Rural Appraisal SA: Social Assessment TFS: Township Forestry Station Social Assessment Report for Hunan Forest Restoration and Development Project ABSTRACT ................................................................................................................... 1 1. PROJECT BACKGROUND.................................................................................... 10 1.1 Project Background ........................................................................................ 10 1.2 Project Objectives .......................................................................................... 10 1.3 Project Components ....................................................................................... 10 2. PROCESS AND METHODS OF SA ...................................................................... 11 2.1 Process .......................................................................................................... -
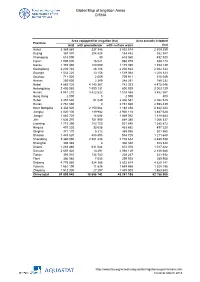
Global Map of Irrigation Areas CHINA
Global Map of Irrigation Areas CHINA Area equipped for irrigation (ha) Area actually irrigated Province total with groundwater with surface water (ha) Anhui 3 369 860 337 346 3 032 514 2 309 259 Beijing 367 870 204 428 163 442 352 387 Chongqing 618 090 30 618 060 432 520 Fujian 1 005 000 16 021 988 979 938 174 Gansu 1 355 480 180 090 1 175 390 1 153 139 Guangdong 2 230 740 28 106 2 202 634 2 042 344 Guangxi 1 532 220 13 156 1 519 064 1 208 323 Guizhou 711 920 2 009 709 911 515 049 Hainan 250 600 2 349 248 251 189 232 Hebei 4 885 720 4 143 367 742 353 4 475 046 Heilongjiang 2 400 060 1 599 131 800 929 2 003 129 Henan 4 941 210 3 422 622 1 518 588 3 862 567 Hong Kong 2 000 0 2 000 800 Hubei 2 457 630 51 049 2 406 581 2 082 525 Hunan 2 761 660 0 2 761 660 2 598 439 Inner Mongolia 3 332 520 2 150 064 1 182 456 2 842 223 Jiangsu 4 020 100 119 982 3 900 118 3 487 628 Jiangxi 1 883 720 14 688 1 869 032 1 818 684 Jilin 1 636 370 751 990 884 380 1 066 337 Liaoning 1 715 390 783 750 931 640 1 385 872 Ningxia 497 220 33 538 463 682 497 220 Qinghai 371 170 5 212 365 958 301 560 Shaanxi 1 443 620 488 895 954 725 1 211 648 Shandong 5 360 090 2 581 448 2 778 642 4 485 538 Shanghai 308 340 0 308 340 308 340 Shanxi 1 283 460 611 084 672 376 1 017 422 Sichuan 2 607 420 13 291 2 594 129 2 140 680 Tianjin 393 010 134 743 258 267 321 932 Tibet 306 980 7 055 299 925 289 908 Xinjiang 4 776 980 924 366 3 852 614 4 629 141 Yunnan 1 561 190 11 635 1 549 555 1 328 186 Zhejiang 1 512 300 27 297 1 485 003 1 463 653 China total 61 899 940 18 658 742 43 241 198 52 -

48443-002: Hunan Xiangjiang River
Resettlement Plan May 2018 People’s Republic of China: Hunan Xiangjiang River Watershed Existing Solid Waste Comprehensive Treatment Project --- Changning Subproject Prepared by The People’s Government of Changning for the Asian Development Bank. CURRENCY EQUIVALENTS (as of 30 April 2018) Currency unit – yuan (CNY) CNY1.00 = $0.15 8 $1.00 = CNY6.334 ABBREVIATIONS AAOV – average annual output value ADB – Asian Development Bank DI – design institute EA – executing agency FSR – feasibility study report HD – House demolition HPG – Hunan Provincial Government IA – implementing agency LA – land acquisition LAR – land acquisition and resettlement LRB – Land and Resources Administration Bureau RP – resettlement plan SES – social and economic survey SPS – ADB’s Safeguard Policy Statement (2009) WEIGHTS AND MEASURES m2 – square meter mu – 1 mu is equal to 666.7 m 2 km – kilometer NOTE In this report, "$" refers to United States dollars. This resettlement plan is a document of the borrower. The views expressed herein do not necessarily represent those of ADB's Board of Directors, Management, or staff, and may be preliminary in nature. Your attention is directed to the “terms of use” section of this website. In preparing any country program or strategy, financing any project, or by making any designation of or reference to a particular territory or geographic area in this document, the Asian Development Bank does not intend to make any judgments as to the legal or other status of any territory or area. Table of Contents EXECUTIVE SUMMARY ....................................................................................................................... -

Resettlement Plan: Zixing Subproject
Resettlement Plan May 2018 People’s Republic of China: Hunan Xiangjiang River Watershed Existing Solid Waste Comprehensive Treatment Project --- Zixing Subproject Prepared by the People’s Government of Zixing City for the Asian Development Bank. CURRENCY EQUIVALENTS (as of 30 April 2018) Currency unit – yuan (CNY) CNY1.00 = $0.15 8 $1.00 = CNY6.334 ABBREVIATIONS AAOV – average annual output value ADB – Asian Development Bank DI – design institute EA – executing agency FS R – feasibility study report HD – House demolition IA – implementing agency LA – land acquisition LAR – land acquisition and resettlement LRB – Land and Resources Admi nistration Bureau NDRC – National Development and Reform Commission RP – resettlement plan SES – social and economic survey SPS – ADB’s Safeguard Policy Statement (2009) WEIGHTS AND MEASURES m2 – square meter mu – equal to 666.7 m 2 km – kilometer NOTE In this report, "$" refers to United States dollars. This resettlement plan is a document of the borrower. The views expressed herein do not necessarily represent those of ADB's Board of Directors, Management, or staff, and may be preliminary in nature. Your attention is directed to the “terms of use” section of this website. In preparing any country program or strategy, financing any project, or by making any designation of or reference to a particular territory or geographic area in this document, the Asian Development Bank does not intend to make any judgments as to the legal or other status of any territory or area. Table of Contents EXECUTIVE SUMMARY ............................................................................................. I 1. PROJECT OVERVIEW .................................................................................... 1 1.1. PROJECT BACKGROUND ............................................................................. 1 1.2 PROJECT IMPACT SCOPE AND LOCATION ............................................... -

Chinese Bankers Survey 2016
www.pwchk.com Chinese Bankers Survey 2016 Executive summary February 2017 Preface We are pleased to present the Chinese Bankers Most bankers in the survey believe that the top Survey 2016 report, prepared jointly by the China priority in addressing risk is "to improve cash Banking Association (CBA) and PwC. Now in its collection, accelerate write-offs and disposal of eighth year, the report keeps track of developments collaterals". While increasing NPL disposal efforts in China’s banking sector from the perspective of a through conventional means such as write-offs, group of bankers. collections and transfers, Chinese banks are also actively exploring a diversified range of alternatives, This year’s survey dug deep into the core issues including NPL securitisation, cooperating with the while maintaining a broad scope. Dr. BA Shusong, big four asset management companies (AMCs), and the project leader, together with the project team, transfer of income beneficial rights. There is a interviewed 15 senior bankers to get their insights section in this report summarising bankers’ views into the sector. Seven of the 15 bankers are at C- on the major challenges regarding disposal of NPLs. suite level (director, vice president or above). These interviews complemented the online survey With China’s economic structural reform at a critical covering 31 provinces in mainland China (excluding juncture, Chinese banks are expected to play a more Hong Kong, Macau and Taiwan). With a total of active role in fulfilling social responsibilities by 1,794 valid responses collected, the survey sampling serving the real economy. With regard to economic process has taken into account participants’ restructuring, most bankers believe that “upgrading geographical regions, grades, types of financial and transforming manufacturing sector, and institution, and if the institution is listed on stock exploration of new markets" brings many exchange(s) or privately held. -

Resource-Based City Type and Reforming Strategy Discussion and Research
2016 4th International Conference on Advances in Social Science, Humanities, and Management (ASSHM 2016) ISBN: 978-1-60595-412-7 Resource-based City Type and Reforming Strategy Discussion and Research Junwei Xing1 Abstract The resource-based city can divides according to the Israeli resource type and development phase two big standards. The resource-based city economic transformation strategy first should from the macroscopic level, seek for the regional economic development the new superiority, next should act according to the new regional development favorable condition that establishes the pattern of industrial transformation from the microscopic level. Keywords. Resource-based City; Type; Reforming Strategy. 1. INTRODUCTION The resource-based city is our country important foundation energy and raw material supplying place, development display excessively significant role to our country national economy. But as a result of the non-renewability of finiteness and resources of resource, the resource-based city sooner or later must face the issue of economic transformation, otherwise possibly enters the winter. At present, a considerable number of resource-based cities in China have been facing a series of economic and social problems such as resource depletion, economic recession, environmental degradation, unemployment and the increase of the poor population. These resource-based cities have become the problem areas in which regional contradictions are concentrated. Therefore, how to guide these cities to successfully transform and achieve sustainable development has become an important topic of concern for both academia and government. 2. Classification of Resource-based Cities Resource based cities are mainly built up by the development of resources. Although the overall pattern of urban growth and development is similar, because of the differences of the economic development factors such as natural resources, geographical conditions and other economic development factors, the difference between different resource cities is very large. -

Improved Chaotic Quantum-Behaved Particle Swarm Optimization Algorithm for Fuzzy Neural Network and Its Application
Hindawi Mathematical Problems in Engineering Volume 2020, Article ID 9464593, 11 pages https://doi.org/10.1155/2020/9464593 Research Article Improved Chaotic Quantum-Behaved Particle Swarm Optimization Algorithm for Fuzzy Neural Network and Its Application Yuexi Peng ,1 Kejun Lei ,2 Xi Yang,2 and Jinzhang Peng 3, 1School of Physics and Electronics, Central South University, Changsha 410083, China 2College of Information Science and Engineering, Jishou University, Jishou 416000, China 3College of Physics and Mechatronics Engineering, Jishou University, Jishou 416000, China Correspondence should be addressed to Kejun Lei; [email protected] Received 7 February 2020; Accepted 5 March 2020; Published 28 March 2020 Guest Editor: Kehui Sun Copyright © 2020 Yuexi Peng et al. 0is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Traditional fuzzy neural network has certain drawbacks such as long computation time, slow convergence rate, and premature convergence. To overcome these disadvantages, an improved quantum-behaved particle swarm optimization algorithm is proposed as the learning algorithm. In this algorithm, a new chaotic search is introduced, and benchmark function experiments prove it outperforms the other five existing algorithms. Finally, the proposed algorithm is presented as the learning algorithm for Takagi–Sugeno fuzzy neural network to form a new neural network, and it is utilized in the water quality evaluation of Dongjiang Lake of Hunan province. Simulation results demonstrated the effectiveness of the new neural network. 1. Introduction is a complex high-dimensional problem, PSO algorithm has the disadvantage of premature convergence [16–19]. -

Primulina Zixingensis (Gesneriaceae), a New Species from Hunan, China
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/341391934 Primulina Zixingensis (Gesneriaceae), a New Species from Hunan, China Article in Annales Botanici Fennici · May 2020 DOI: 10.5735/085.057.0107 CITATIONS READS 2 69 4 authors, including: Meizhen Xu Lihua Yang Chinese Academy of Sciences Chinese Academy of Sciences 19 PUBLICATIONS 9 CITATIONS 24 PUBLICATIONS 66 CITATIONS SEE PROFILE SEE PROFILE All content following this page was uploaded by Lihua Yang on 04 September 2020. The user has requested enhancement of the downloaded file. Primulina zixingensis (Gesneriaceae), a New Species from Hunan, China Authors: Pan, Bo, Xu, Mei-Zhen, Tang, Wen-Xiu, and Yang, Li-Hua Source: Annales Botanici Fennici, 57(1-3) : 55-59 Published By: Finnish Zoological and Botanical Publishing Board URL: https://doi.org/10.5735/085.057.0107 BioOne Complete (complete.BioOne.org) is a full-text database of 200 subscribed and open-access titles in the biological, ecological, and environmental sciences published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Complete website, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/terms-of-use. Usage of BioOne Complete content is strictly limited to personal, educational, and non - commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. -

World Bank Document
Document of The World Bank FOR OFFICIAL USE ONLY Public Disclosure Authorized Report No: 67628-CN PROJECT APPRAISAL DOCUMENT ON THREE PROPOSED LOANS IN A TOTAL AMOUNT OF Public Disclosure Authorized US$150 MILLION (US$50 MILLION EACH TO FINANCE PROJECT ACTIVITIES IN GUANGDONG, HUNAN AND GANSU PROVINCES) TO THE PEOPLE’S REPUBLIC OF CHINA FOR THE Public Disclosure Authorized INTEGRATED ECONOMIC DEVELOPMENT OF SMALL TOWNS PROJECT April 23, 2012 China and Mongolia Sustainable Development Unit Sustainable Development Department East Asia and Pacific Region Public Disclosure Authorized This document has a restricted distribution and may be used by recipients only in the performance of their official duties. Its contents may not otherwise be disclosed without World Bank authorization. CURRENCY EQUIVALENTS (Exchange Rate Effective: December 9, 2011) Currency Unit = Renminbi (RMB) Yuan RMB Yuan 1.0 = US$ 0.156 US$ 1.0 = RMB Yuan 6.39 FISCAL YEAR July 1 – June 30 ABBREVIATIONS AND ACRONYMS AWP Annual Work Plan CAS Country Assistance Strategy CEA Cost-Effectiveness Analysis CHM Complain Handle Mechanism CNAO Chinese National Audit Office CPMO County Project Management Office CPS Country Partnership Strategy CQ Consultant Qualifications CWG Consulting Working Group DA Designated Account DIs Design Institutes EA Environmental Assessment ECOP Environmental Code of Practice EIA Environmental Impact Assessment EMDP Ethnic Minority Development Plan EMP Environmental Management Plan ERR Economic Rate of Return FM Financial Management FMM Financial Management -

The Efficiency of Primary Health Care Institutions in the Counties
International Journal of Environmental Research and Public Health Article The Efficiency of Primary Health Care Institutions in the Counties of Hunan Province, China: Data from 2009 to 2017 Kaili Zhong 1, Lv Chen 1,*, Sixiang Cheng 1, Hongjun Chen 2 and Fei Long 2 1 Department of Social Medicine and Health Management, Xiangya School of Public Health, Central South University, Changsha 410078, China; [email protected] (K.Z.); [email protected] (S.C.) 2 Department of Primary Health Care, Health Commission of Hunan Province, Changsha 410078, China; [email protected] (H.C.); [email protected] (F.L.) * Correspondence: [email protected] Received: 24 December 2019; Accepted: 7 March 2020; Published: 9 March 2020 Abstract: This study aimed to estimate the efficiency and its influencing factors of Primary Health Care Institutions (PHCIs) in counties in Hunan Province, China, and put forward feasible suggestions for improving the efficiency of PHCIs in Hunan Province. We applied the Input-Oriented Data Envelopment Analysis (DEA) method and the Malmquist Index Model to estimate the efficiency of PHCIs in 86 counties in Hunan Province from 2009 to 2017. Then, the Tobit model was used to estimate the factors that influence the efficiency of PHCIs. Since the implementation of the new health-care reform in 2009, the number of health resources in PHCIs in Hunan Province has increased significantly, but most counties’ PHCIs remain inefficient. The efficiency of PHCIs is mainly affected by the total population, city level, the proportion of health technicians and the proportion of beds, but the changes in per capita GDP have not yet played a significant role in influencing efficiency. -

Vertical Facility List
Facility List The Walt Disney Company is committed to fostering safe, inclusive and respectful workplaces wherever Disney-branded products are manufactured. Numerous measures in support of this commitment are in place, including increased transparency. To that end, we have published this list of the roughly 7,600 facilities in over 70 countries that manufacture Disney-branded products sold, distributed or used in our own retail businesses such as The Disney Stores and Theme Parks, as well as those used in our internal operations. Our goal in releasing this information is to foster collaboration with industry peers, governments, non- governmental organizations and others interested in improving working conditions. Under our International Labor Standards (ILS) Program, facilities that manufacture products or components incorporating Disney intellectual properties must be declared to Disney and receive prior authorization to manufacture. The list below includes the names and addresses of facilities disclosed to us by vendors under the requirements of Disney’s ILS Program for our vertical business, which includes our own retail businesses and internal operations. The list does not include the facilities used only by licensees of The Walt Disney Company or its affiliates that source, manufacture and sell consumer products by and through independent entities. Disney’s vertical business comprises a wide range of product categories including apparel, toys, electronics, food, home goods, personal care, books and others. As a result, the number of facilities involved in the production of Disney-branded products may be larger than for companies that operate in only one or a limited number of product categories. In addition, because we require vendors to disclose any facility where Disney intellectual property is present as part of the manufacturing process, the list includes facilities that may extend beyond finished goods manufacturers or final assembly locations. -
January to December 1999
CHINA Death Penalty log: January to December 1999 The table overleaf lists the following (from left to right): the sources and dates of the reports (when known); the date of the reported death sentences (DS) or executions (EX) (when known); the province or city where the sentences were imposed or carried out (when known); the names (when known) or numbers of the alleged offenders and further details (when known); the alleged criminal offences for which the death sentences were imposed (when known); the number of death sentences imposed without stay of execution; the number of executions actually carried out; the number of death sentences imposed with a two-year stay of execution (2y); other information when available, including the number of death sentences, executions and two-year reprieve cases for each month. Abbreviations used in the table: DS number of death sentences pronounced without a stay of execution EX number of executions reported to have been carried out 2y number of death sentences with a 2-year reprieve (stay of execution) (F) Female Source of Report (unless otherwise given) SWB BBC Summary of World Broadcasts (U.K. publication) SCMP South China Morning Post (Hong Kong daily newspaper) FBIS Foreign Broadcast Information Service (U.S. publication) AFP Agence France Presse AP Associated Press Xinhua New China News Agency (Chinese state news agency) VOA Voice of America CNA China News Analysis ETIC East Turkistan Information Center PBS People's Broadcasting Station (Chinese state radio service) Alleged Crime A Arson