E26. Selective Oestrogen Enzyme Modulators (Seems) and Their Effect
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
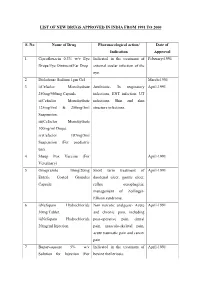
List of New Drugs Approved in India from 1991 to 2000
LIST OF NEW DRUGS APPROVED IN INDIA FROM 1991 TO 2000 S. No Name of Drug Pharmacological action/ Date of Indication Approval 1 Ciprofloxacin 0.3% w/v Eye Indicated in the treatment of February-1991 Drops/Eye Ointment/Ear Drop external ocular infection of the eye. 2 Diclofenac Sodium 1gm Gel March-1991 3 i)Cefaclor Monohydrate Antibiotic- In respiratory April-1991 250mg/500mg Capsule. infections, ENT infection, UT ii)Cefaclor Monohydrate infections, Skin and skin 125mg/5ml & 250mg/5ml structure infections. Suspension. iii)Cefaclor Monohydrate 100mg/ml Drops. iv)Cefaclor 187mg/5ml Suspension (For paediatric use). 4 Sheep Pox Vaccine (For April-1991 Veterinary) 5 Omeprazole 10mg/20mg Short term treatment of April-1991 Enteric Coated Granules duodenal ulcer, gastric ulcer, Capsule reflux oesophagitis, management of Zollinger- Ellison syndrome. 6 i)Nefopam Hydrochloride Non narcotic analgesic- Acute April-1991 30mg Tablet. and chronic pain, including ii)Nefopam Hydrochloride post-operative pain, dental 20mg/ml Injection. pain, musculo-skeletal pain, acute traumatic pain and cancer pain. 7 Buparvaquone 5% w/v Indicated in the treatment of April-1991 Solution for Injection (For bovine theileriosis. Veterinary) 8 i)Kitotifen Fumerate 1mg Anti asthmatic drug- Indicated May-1991 Tablet in prophylactic treatment of ii)Kitotifen Fumerate Syrup bronchial asthma, symptomatic iii)Ketotifen Fumerate Nasal improvement of allergic Drops conditions including rhinitis and conjunctivitis. 9 i)Pefloxacin Mesylate Antibacterial- In the treatment May-1991 Dihydrate 400mg Film Coated of severe infection in adults Tablet caused by sensitive ii)Pefloxacin Mesylate microorganism (gram -ve Dihydrate 400mg/5ml Injection pathogens and staphylococci). iii)Pefloxacin Mesylate Dihydrate 400mg I.V Bottles of 100ml/200ml 10 Ofloxacin 100mg/50ml & Indicated in RTI, UTI, May-1991 200mg/100ml vial Infusion gynaecological infection, skin/soft lesion infection. -

Classification and Pharmacology of Progestins
Maturitas 46S1 (2003) S7–S16 Classification and pharmacology of progestins Adolf E. Schindler a,∗, Carlo Campagnoli b, René Druckmann c, Johannes Huber d, Jorge R. Pasqualini e, Karl W. Schweppe f, Jos H. H. Thijssen g a Institut für Medizinische Forschung und Fortbildung, Universitätsklinikum, Hufelandstr. 55, Essen 45147, Germany b Ospedale Ginecologico St. Anna, Corso Spezia 60, 10126 Torino, Italy c Ameno-Menopause-Center, 12, Rue de France, 06000 Nice, France d Abt. für Gynäkologische Endokrinologie, AKH Wien, Währingergürtel 18-20, 1090 Wien, Austria e Institute de Puériculture26, Boulevard Brune, 75014 Paris, France f Abt. für Gynäkologie und Geburtshilfe, Ammerland Klinik, Langestr.38, 26622 Westerstede, Germany g Department of Endocrinology, Universitair Medisch Centrum Utrecht, P.O. Box 85090, 3508 AB Utrecht, The Netherlands Abstract Besides the natural progestin, progesterone, there are different classes of progestins, such as retroprogesterone (i.e. dydroges- terone), progesterone derivatives (i.e. medrogestone) 17␣-hydroxyprogesterone derivatives (i.e. chlormadinone acetate, cypro- terone acetate, medroxyprogesterone acetate, megestrol acetate), 19-norprogesterone derivatives (i.e. nomegestrol, promege- stone, trimegestone, nesterone), 19-nortestosterone derivatives norethisterone (NET), lynestrenol, levonorgestrel, desogestrel, gestodene, norgestimate, dienogest) and spironolactone derivatives (i.e. drospirenone). Some of the synthetic progestins are prodrugs, which need to be metabolized to become active compounds. Besides -

Northern Ireland Prescription Code Book Drugs Section September 2021
Family Practitioner Services, Pharmaceutical Services, 2 Franklin Street, Belfast BT2 8DQ Telephone No. 028 9053 5613, Fax No. 028 9053 2963 Northern Ireland Prescription Code Book Drugs section September 2021 Page 1 of 587 Effective for prescriptions dispensed September 2021 BSO Code dm+d Pack Description Code Number Special ZD Of Container 38960 4SURE beta-ketone testing strips 4S-810-4183401-001 (Nipro Diagnostics (UK) Ltd) 10 strip strips 38961 4SURE testing strips (Nipro Diagnostics (UK) Ltd) 50 strip strips 4144 AAA 1.5mg/dose sore throat spray (Manx Healthcare Ltd) 60 dose [BSO pack = 1] BSO Unit Of Measure Code No. devices dispensed 70598 (DT) Abacavir 600mg / Lamivudine 300mg tablets 30 tablet tablets 9154 Abasaglar 100units/ml solution for injection 3ml cartridges (Eli Lilly and Company Ltd) 5 cartridge cartridges ZD 9038 Abasaglar KwikPen 100units/ml solution for injection 3ml pre-filled pens (Eli Lilly and Company Ltd) 5 pre- injections ZD filled disposable injection 70088 (DT) Abatacept 125mg/1ml solution for injection pre-filled disposable devices 4 pre-filled disposable injections ZD injection 70089 (DT) Abatacept 125mg/1ml solution for injection pre-filled syringes 4 pre-filled disposable injection injections ZD 70144 Abelcet 100mg/20ml concentrate for suspension for infusion vials (Teva UK Ltd) 10 vial vials Hospital Only 3303 Abidec Multivitamin drops (Omega Pharma Ltd) 25 ml mls 4043 Abilify 10mg orodispersible tablets (Otsuka Pharmaceuticals (U.K.) Ltd) 28 tablet 4 x 7 tablets tablets 3006 Abilify 10mg tablets (Otsuka -

209627Orig1s000
CENTER FOR DRUG EVALUATION AND RESEARCH APPLICATION NUMBER: 209627Orig1s000 MULTI-DISCIPLINE REVIEW Summary Review Office Director Cross Discipline Team Leader Review Clinical Review Non-Clinical Review Statistical Review Clinical Pharmacology Review Reviewers of Multi-Disciplinary Review and Evaluation SECTIONS OFFICE/ AUTHORED/ ACKNOWLEDGED/ DISCIPLINE REVIEWER DIVISION APPROVED Mark Seggel, Ph.D. OPQ/ONDP/DNDP2 Authored: Section 4.2 Digitally signed by Mark R. Seggel -S CMC Lead DN: c=US, o=U.S. Government, ou=HHS, ou=FDA, ou=People, cn=Mark R. Signature: Mark R. Seggel -S Seggel -S, 0.9.2342.19200300.100.1.1=1300071539 Date: 2018.08.08 16:29:15 -04'00' Frederic Moulin, DVM, PhD OND/ODE3/DBRUP Authored: Section 5 Pharmacology/ Digitally signed by Frederic Moulin -S Toxicology DN: c=US, o=U.S. Government, ou=HHS, ou=FDA, ou=People, Reviewer Signature: Frederic Moulin -S 0.9.2342.19200300.100.1.1=2001708658, cn=Frederic Moulin -S Date: 2018.08.08 15:26:57 -04'00' Kimberly Hatfield, PhD OND/ODE3/DBRUP Approved: Section 5 Pharmacology/ Toxicology Digitally signed by Kimberly P. Hatfield -S DN: c=US, o=U.S. Government, ou=HHS, ou=FDA, ou=People, Team Leader Signature: Kimberly P. Hatfield -S 0.9.2342.19200300.100.1.1=1300387215, cn=Kimberly P. Hatfield -S Date: 2018.08.08 14:56:10 -04'00' Li Li, Ph.D. OCP/DCP3 Authored: Sections 6 and 17.3 Clinical Pharmacology Dig ta ly signed by Li Li S DN c=US o=U S Government ou=HHS ou=FDA ou=People Reviewer cn=Li Li S Signature: Li Li -S 0 9 2342 19200300 100 1 1=20005 08577 Date 2018 08 08 15 39 23 04'00' Doanh Tran, Ph.D. -

A Review of Mastalgia in Patients with Fibrocystic Breast Changes and the Non-Surgical Treatment Options
REVIEW ARTICLE A Review of Mastalgia in Patients with Fibrocystic Breast Changes and the Non-Surgical Treatment Options Khalid Rida Murshid FRCS(C), FACS Department of Surgery, College of Medicine, Taibah University, Al Madinah Al Munawwarah, Kingdom of Saudi Arabia Abstract Objectives The objectives of this study are to review fibrocystic changes of the breast, their causal and associated factors and their correlation to mastalgia, and then to review the available treatment options for mastalgia (caused by fibrocystic changes) short of surgery. Methods The author reviews all the articles obtained from a PubMed research on mastalgia and fibrocystic changes of the breast, published in English over the last 14 years. Results Fibrocystic changes of the breast are common and can be considered as a normal phase of breast development. These changes are sometimes asymptomatic; however, when painful, patients would seek medical advice. Lifestyle changes and the avoidance of certain dietary elements as well as the use of some non-pharmacological agents have shown some beneficiary effects. In severe cases, stronger pharmacological and hormonal agents are resorted to being more effective but are associated with greater side effects. Conclusion Fibrocystic changes of the breast are common and should not be considered a disease. When painful, reassurance and non-pharmacological measures should be used first as a treatment. Stronger pharmacological and hormonal agents hold more serious side effects. Some of these remedies are supported by good clinical evidence, while others are not. The ideal treatment for mastalgia caused by fibrocystic changes is to be identified by sound recent randomized controlled clinical studies on simple remedies before being performed on stronger ones. -

Part I Biopharmaceuticals
1 Part I Biopharmaceuticals Translational Medicine: Molecular Pharmacology and Drug Discovery First Edition. Edited by Robert A. Meyers. © 2018 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2018 by Wiley-VCH Verlag GmbH & Co. KGaA. 3 1 Analogs and Antagonists of Male Sex Hormones Robert W. Brueggemeier The Ohio State University, Division of Medicinal Chemistry and Pharmacognosy, College of Pharmacy, Columbus, Ohio 43210, USA 1Introduction6 2 Historical 6 3 Endogenous Male Sex Hormones 7 3.1 Occurrence and Physiological Roles 7 3.2 Biosynthesis 8 3.3 Absorption and Distribution 12 3.4 Metabolism 13 3.4.1 Reductive Metabolism 14 3.4.2 Oxidative Metabolism 17 3.5 Mechanism of Action 19 4 Synthetic Androgens 24 4.1 Current Drugs on the Market 24 4.2 Therapeutic Uses and Bioassays 25 4.3 Structure–Activity Relationships for Steroidal Androgens 26 4.3.1 Early Modifications 26 4.3.2 Methylated Derivatives 26 4.3.3 Ester Derivatives 27 4.3.4 Halo Derivatives 27 4.3.5 Other Androgen Derivatives 28 4.3.6 Summary of Structure–Activity Relationships of Steroidal Androgens 28 4.4 Nonsteroidal Androgens, Selective Androgen Receptor Modulators (SARMs) 30 4.5 Absorption, Distribution, and Metabolism 31 4.6 Toxicities 32 Translational Medicine: Molecular Pharmacology and Drug Discovery First Edition. Edited by Robert A. Meyers. © 2018 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2018 by Wiley-VCH Verlag GmbH & Co. KGaA. 4 Analogs and Antagonists of Male Sex Hormones 5 Anabolic Agents 32 5.1 Current Drugs on the Market 32 5.2 Therapeutic Uses and Bioassays -

Etats Rapides
List of European Pharmacopoeia Reference Standards Effective from 2015/12/24 Order Reference Standard Batch n° Quantity Sale Information Monograph Leaflet Storage Price Code per vial Unit Y0001756 Exemestane for system suitability 1 10 mg 1 2766 Yes +5°C ± 3°C 79 ! Y0001561 Abacavir sulfate 1 20 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0001552 Abacavir for peak identification 1 10 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0001551 Abacavir for system suitability 1 10 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0000055 Acamprosate calcium - reference spectrum 1 n/a 1 1585 79 ! Y0000116 Acamprosate impurity A 1 50 mg 1 3-aminopropane-1-sulphonic acid 1585 Yes +5°C ± 3°C 79 ! Y0000500 Acarbose 3 100 mg 1 See leaflet ; Batch 2 is valid until 31 August 2015 2089 Yes +5°C ± 3°C 79 ! Y0000354 Acarbose for identification 1 10 mg 1 2089 Yes +5°C ± 3°C 79 ! Y0000427 Acarbose for peak identification 3 20 mg 1 Batch 2 is valid until 31 January 2015 2089 Yes +5°C ± 3°C 79 ! A0040000 Acebutolol hydrochloride 1 50 mg 1 0871 Yes +5°C ± 3°C 79 ! Y0000359 Acebutolol impurity B 2 10 mg 1 -[3-acetyl-4-[(2RS)-2-hydroxy-3-[(1-methylethyl)amino] propoxy]phenyl] 0871 Yes +5°C ± 3°C 79 ! acetamide (diacetolol) Y0000127 Acebutolol impurity C 1 20 mg 1 N-(3-acetyl-4-hydroxyphenyl)butanamide 0871 Yes +5°C ± 3°C 79 ! Y0000128 Acebutolol impurity I 2 0.004 mg 1 N-[3-acetyl-4-[(2RS)-3-(ethylamino)-2-hydroxypropoxy]phenyl] 0871 Yes +5°C ± 3°C 79 ! butanamide Y0000056 Aceclofenac - reference spectrum 1 n/a 1 1281 79 ! Y0000085 Aceclofenac impurity F 2 15 mg 1 benzyl[[[2-[(2,6-dichlorophenyl)amino]phenyl]acetyl]oxy]acetate -

Giving Another Chance to Mifepristone in Pharmacotherapy for Aggressive Meningiomas—A Likely Synergism with Hydroxyurea?
Curr Probl Cancer 40 (2016) 229–243 Contents lists available at ScienceDirect Curr Probl Cancer journal homepage: www.elsevier.com/locate/cpcancer Giving another chance to mifepristone in pharmacotherapy for aggressive meningiomas—A likely synergism with hydroxyurea? İlhan Elmaci, MDa, Meric A. Altinoz, MDb,*, Aydin Sav, MDc, Zeliha Yazici, PhDd, Aysel Ozpinar, PhDe Introduction Meningiomas: An underestimated health problem Meningiomas are the most frequently reported intracranial tumors, accounting for approx- imately one-fourth of all reported primary brain neoplasms.1 They are benign in approximately 90% of the cases and the remaining cases are either borderline or atypical (World Health Organization [WHO] grade II) or malignant (WHO grade III).1 In the United States, the incidence rates with similar age standardization estimated from figures provided by the Central Brain Tumor Registrywere1.8formenand4.2per100,000forwomenin2006.1 Increasing incidence rates of meningiomas have been reported from several industrialized countries since the early 1980s. But recent studies revealed that real meningioma incidence is even higher.1 As meningiomas are mostly benign, they are not covered by most cancer registries.1 Nevertheless, in Finland, as in other Nordic countries, all surgeons and pathologists are obliged to report all tumors of the central nervous system, both malignant and benign, to the cancer registry.1 To reveal the real incidence of Grant/Financial Support: none. This is original work and is not under consideration for publication elsewhere. This submission is for the special issue being compiled by Beata Holkova on the topic Amylodosis. a Department of Neurosurgery, Memorial Hospital, Istanbul, Turkey b Department of Immunology, Experimental Medical Research Institute/DETAE, Istanbul University, Istanbul, Turkey c Nisantasi Neuropathology Group, Istanbul, Turkey d Department of Pharmacology, Cerrahpasa Faculty of Medicine, Istanbul University, Turkey e Department of Biochemistry, Acibadem University, Istanbul, Turkey * Corresponding author: Meric A. -

Progestogens and Venous Thromboembolism Among Postmenopausal Women Using Hormone Therapy. Marianne Canonico, Geneviève Plu-Bureau, Pierre-Yves Scarabin
Progestogens and venous thromboembolism among postmenopausal women using hormone therapy. Marianne Canonico, Geneviève Plu-Bureau, Pierre-Yves Scarabin To cite this version: Marianne Canonico, Geneviève Plu-Bureau, Pierre-Yves Scarabin. Progestogens and venous throm- boembolism among postmenopausal women using hormone therapy.. Maturitas, Elsevier, 2011, 70 (4), pp.354-60. 10.1016/j.maturitas.2011.10.002. inserm-01148705 HAL Id: inserm-01148705 https://www.hal.inserm.fr/inserm-01148705 Submitted on 5 May 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Progestogens and VTE Finale version Progestogens and venous thromboembolism among postmenopausal women using hormone therapy Marianne Canonico1,2, Geneviève Plu-Bureau1,3 and Pierre-Yves Scarabin1,2 1 Centre for Research in Epidemiology and Population Health, U1018, Hormones and Cardiovascular Disease 2 University Paris-Sud, UMR-S 1018, Villejuif, France 3 University Paris Descartes and Hôtel-Dieu Hospital, Paris, France Adresse: 16 av. Paul Vaillant Couturier 94807 Villejuif Cedex Tel: +33 1 45 59 51 66 Fax: +33 1 45 59 51 70 Corresponding author: Marianne Canonico ([email protected]) 1/21 Progestogens and VTE Finale version Abstract Hormone therapy (HT) is the most effective treatment for correcting menopausal symptoms after menopause. -

Minutes of PRAC Meeting on 09-12 July 2018
6 September 2018 EMA/PRAC/576790/2018 Inspections, Human Medicines Pharmacovigilance and Committees Division Pharmacovigilance Risk Assessment Committee (PRAC) Minutes of the meeting on 09-12 July 2018 Chair: June Raine – Vice-Chair: Almath Spooner Health and safety information In accordance with the Agency’s health and safety policy, delegates were briefed on health, safety and emergency information and procedures prior to the start of the meeting. Disclaimers Some of the information contained in the minutes is considered commercially confidential or sensitive and therefore not disclosed. With regard to intended therapeutic indications or procedure scope listed against products, it must be noted that these may not reflect the full wording proposed by applicants and may also change during the course of the review. Additional details on some of these procedures will be published in the PRAC meeting highlights once the procedures are finalised. Of note, the minutes are a working document primarily designed for PRAC members and the work the Committee undertakes. Note on access to documents Some documents mentioned in the minutes cannot be released at present following a request for access to documents within the framework of Regulation (EC) No 1049/2001 as they are subject to on- going procedures for which a final decision has not yet been adopted. They will become public when adopted or considered public according to the principles stated in the Agency policy on access to documents (EMA/127362/2006, Rev. 1). 30 Churchill Place ● Canary Wharf ● London E14 5EU ● United Kingdom Telephone +44 (0)20 3660 6000 Facsimile +44 (0)20 3660 5555 Send a question via our website www.ema.europa.eu/contact An agency of the European Union © European Medicines Agency, 2018. -

Rominder P.S. Suri, Phd, PE Robert F. Chimchirian & Sandhya Meduri Department of Civil & Environmental Engineering Villa
Emerging Contaminants and Water Supply – Speaker Abstracts Emerging Contaminants and Water Supply – Speaker Abstracts PRESENCE OF ESTROGEN HORMONES AND ANTIBIOTICS IN detection limits for all 12 estrogen compounds ranged from 0.1 ng L-1 to THE ENVIRONMENT 10 ng L-1 using a sample size of 1 L. Eight antibiotics and venlaflaxine Rominder P.S. Suri, PhD, PE were detected in the effluent with concentrations that ranged from 50 to Robert F. Chimchirian & Sandhya Meduri 2,930 ng L-1. Department of Civil & Environmental Engineering Twenty-one streams in Chester County, southeast Pennsylvania, were Villanova University, Villanova PA 19085 also sampled for aqueous concentrations of the above hormones and 610-519-7631 [email protected] antibiotics. Chester County represents a very diverse land use. In addition to concentrated urban developments, there are many In recent years, the detection of trace amount of pharmaceutically active agricultural farms (dairy, cattle, hog, poultry, and mushroom). The compounds (PACs) in the surface water has gained widespread watershed is comprised of about 280,000 acres that eventually drain into attention. The presence of the PACs in the natural water systems causes the Chesapeake Bay via the Susquehanna River. The results showed that ecological problems and possible adverse effects to humans. Two classes there was at least one estrogen detected in every stream sampled and six of PACs, estrogen hormones and antibiotics make their way into the estrogens were detected at three different locations. The most abundant surface waters through effluents from wastewater treatment plants and PAC was estrone, which was detected in all but one stream, with areas where animal manure and biosolids are land applied, amongst concentrations as high as 2.4 ng L-1. -

Synthesis and Regulation of Sex Hormones in Breast Tissue During Pre- and Postmenopause
JOURNAL FÜR MENOPAUSE PASQUALINI JR Synthesis and regulation of sex hormones in breast tissue during pre- and postmenopause Journal für Menopause 2001; 8 (Supplementum 2) (Ausgabe für Schweiz), 10-13 Homepage: www.kup.at/menopause Online-Datenbank mit Autoren- und Stichwortsuche TERIUMTE IM KLIMAKZEITSCHRIFTISCHE ASPEKT FÜR DIAGNOSTISCHE, THERAPEUTISCHE UND PROPHYLAK Indexed in EMBASE/ Krause & Pachernegg GmbH · VERLAG für MEDIZIN und WIRTSCHAFT · A-3003 Gablitz Excerpta Medica SYNTHESIS AND J. R. Pasqualini REGULATION OF SEX HORMONES IN BREAST SYNTHESIS AND REGULATION OF SEX TISSUE DURING HORMONES IN BREAST TISSUE DURING PRE- AND POSTMENOPAUSE PRE- AND POSTMENOPAUSE and the “sulphatase pathway” which INTRODUCTION converts E1S into E1 by the estrone- ESTRONE SULPHATASE sulphatase (EC: 3.1.6.1) [8–10]. The final step of steroidogenesis is the ACTIVITY IN BREAST CANCER Most breast cancers (~ 95 %) are in conversion of the weak E1 to the AND ITS CONTROL their early stage hormone-sensitive, potent biologically active E2 by the where the estrogen estradiol (E2) action of a reductive 17β-hydroxy- plays an important role in the gen- steroid dehydrogenase type 1 activity For many years the endocrine therapy esis and development of this tumour (17β-HSD-1, EC 1.1.1.62) [11–12]. in breast cancer has been mainly by [1, 2]. About two thirds of breast the utilization of antiestrogens (e.g. cancers occur during the postmeno- Quantitative evaluation indicates Nolvadex, tamoxifen citrate) which pausal period when the ovaries have that in human breast tissue E1S “via block the estrogen receptor. ceased to be functional. Despite the sulphatase” is a much more likely low levels of circulating estrogens, precursor for E2 than is androsten- More recently, another endocrine the tissular concentrations of estrone edione “via aromatase” (Fig.