Potential Distribution of the Australian Native Chloris Truncata Based on Modelling Both the Successful and Failed Global Introductions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Kingdom Class Family Scientific Name Common Name I Q a Records
Kingdom Class Family Scientific Name Common Name I Q A Records plants monocots Poaceae Paspalidium rarum C 2/2 plants monocots Poaceae Aristida latifolia feathertop wiregrass C 3/3 plants monocots Poaceae Aristida lazaridis C 1/1 plants monocots Poaceae Astrebla pectinata barley mitchell grass C 1/1 plants monocots Poaceae Cenchrus setigerus Y 1/1 plants monocots Poaceae Echinochloa colona awnless barnyard grass Y 2/2 plants monocots Poaceae Aristida polyclados C 1/1 plants monocots Poaceae Cymbopogon ambiguus lemon grass C 1/1 plants monocots Poaceae Digitaria ctenantha C 1/1 plants monocots Poaceae Enteropogon ramosus C 1/1 plants monocots Poaceae Enneapogon avenaceus C 1/1 plants monocots Poaceae Eragrostis tenellula delicate lovegrass C 2/2 plants monocots Poaceae Urochloa praetervisa C 1/1 plants monocots Poaceae Heteropogon contortus black speargrass C 1/1 plants monocots Poaceae Iseilema membranaceum small flinders grass C 1/1 plants monocots Poaceae Bothriochloa ewartiana desert bluegrass C 2/2 plants monocots Poaceae Brachyachne convergens common native couch C 2/2 plants monocots Poaceae Enneapogon lindleyanus C 3/3 plants monocots Poaceae Enneapogon polyphyllus leafy nineawn C 1/1 plants monocots Poaceae Sporobolus actinocladus katoora grass C 1/1 plants monocots Poaceae Cenchrus pennisetiformis Y 1/1 plants monocots Poaceae Sporobolus australasicus C 1/1 plants monocots Poaceae Eriachne pulchella subsp. dominii C 1/1 plants monocots Poaceae Dichanthium sericeum subsp. humilius C 1/1 plants monocots Poaceae Digitaria divaricatissima var. divaricatissima C 1/1 plants monocots Poaceae Eriachne mucronata forma (Alpha C.E.Hubbard 7882) C 1/1 plants monocots Poaceae Sehima nervosum C 1/1 plants monocots Poaceae Eulalia aurea silky browntop C 2/2 plants monocots Poaceae Chloris virgata feathertop rhodes grass Y 1/1 CODES I - Y indicates that the taxon is introduced to Queensland and has naturalised. -

(Poaceae) and Characterization
EVOLUTION AND DEVELOPMENT OF VEGETATIVE ARCHITECTURE: BROAD SCALE PATTERNS OF BRANCHING ACROSS THE GRASS FAMILY (POACEAE) AND CHARACTERIZATION OF ARCHITECTURAL DEVELOPMENT IN SETARIA VIRIDIS L. P. BEAUV. By MICHAEL P. MALAHY Bachelor of Science in Biology University of Central Oklahoma Edmond, Oklahoma 2006 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the Degree of MASTER OF SCIENCE July, 2012 EVOLUTION AND DEVELOPMENT OF VEGETATIVE ARCHITECTURE: BROAD SCALE PATTERNS OF BRANCHING ACROSS THE GRASS FAMILY (POACEAE) AND CHARACTERIZATION OF ARCHITECTURAL DEVELOPMENT IN WEEDY GREEN MILLET ( SETARIA VIRIDIS L. P. BEAUV.) Thesis Approved: Dr. Andrew Doust Thesis Adviser Dr. Mark Fishbein Dr. Linda Watson Dr. Sheryl A. Tucker Dean of the Graduate College I TABLE OF CONTENTS Chapter Page I. Evolutionary survey of vegetative branching across the grass family (poaceae) ... 1 Introduction ................................................................................................................... 1 Plant Architecture ........................................................................................................ 2 Vascular Plant Morphology ......................................................................................... 3 Grass Morphology ....................................................................................................... 4 Methods ....................................................................................................................... -

MAULES CREEK COAL PROJECT Biodiversity Offset Management Plan
MAULES CREEK COAL PROJECT Biodiversity Offset Management Plan For: Hansen Bailey December 2011 Final Draft PO Box 2474, Carlingford Court 2118 www.cumberlandecology.com.au Report No. 9125RP3 The preparation of this report has been in accordance with the brief provided by the Client and has relied upon the data and results collected at or under the times and conditions specified in the report. All findings, conclusions or recommendations contained within the report are based only on the aforementioned circumstances. The report has been prepared for use by the Client and no responsibility for its use by other parties is accepted by Cumberland Ecology. Revision Date Issued Reviewed by Approved by Date Approved Revision Type 1 David R David R 17/11/11 First draft 2 David R David R 22/11/11 Second draft 3 David R David R 30/11/11 Third draft 4 David R David R 01/12/11 Final Draft Approved by: David Robertson Position: Director Signed: Date: 1 December, 2011 MAULES CREEK COAL PROJECT FINAL DRAFT HANSEN BAILEY 1DECEMBER 2011 Table Of Contents 1 INTRODUCTION 1.1 Purpose 1.1 1.2 Background 1.1 1.2.1 Assessment and Approval Context 1.1 1.2.2 Description of the Impacts 1.2 1.2.3 Consistency with Recovery Plans and Conservation 1.3 Advices 1.2.4 Integration with Local Strategies 1.3 1.3 Aims and Objectives 1.3 1.4 Structure of the Biodiversity Offset Management Plan 1.4 1.5 Terms and Abbreviations 1.5 2 DESCRIPTION OF THE OFFSETS 2.1 General Description 2.1 2.1.1 Northern Offsets 2.1 2.1.2 Eastern and Western Offsets 2.6 2.1.3 Shared Offset Property -

Field Guide to Common Grasses of the Southern Rangelands
Field Guide to Common Grasses of the Southern Rangelands David Blood, Andrew Mitchell, Jane Bradley (Ed) and Jim Addison About this Guide Authors David Blood, Andrew Mitchell, Jane Bradley (Ed) and Jim Addison Acknowledgements The authors would like to thank John Stretch, Wayne Fletcher and PJ Waddell for their advice and support during the preparation of this publication. Thanks also to the pastoralists who provided advice and hospitality during photography and collecting expeditions. Kevin Thiele, Curator of the WA Herbarium for advice regarding sourcing of specimens. And our partners who tolerated periods of absence and distractedness. Copyright ©2015 Rangelands NRM. The Rangelands NRM copyright protects this publication. Except for purposes permitted by the Copyright Act, reproduction by whatever means is prohibited without the prior written permission of Rangelands NRM. www.rangelandswa.com.au, [email protected] ISBN: 978-0-9943781-0-1 - Cited as Rangelands NRM, 2015, Field Guide to Common Grasses of the Southern Rangelands, Rangelands NRM, Perth WA Layout & Design: Swish Design The production of this publication was supported by Rangelands NRM through funding from the Australian Government’s National Landcare Programme. Field Guide to Common Grasses of the Southern Rangelands Wooleen © P. Price Contents Foreword; Grasses of the Southern Rangelands ....................................................................... 4 Introduction ....................................................................................................................................... -

Taxonomy of the Genus Chloris (Gramineae)
Brigham Young University Science Bulletin, Biological Series Volume 19 Number 2 Article 1 3-1974 Taxonomy of the genus Chloris (Gramineae) Dennis E. Anderson Department of Biology, Humboldt State College, Arcata, California 95521 Follow this and additional works at: https://scholarsarchive.byu.edu/byuscib Part of the Anatomy Commons, Botany Commons, Physiology Commons, and the Zoology Commons Recommended Citation Anderson, Dennis E. (1974) "Taxonomy of the genus Chloris (Gramineae)," Brigham Young University Science Bulletin, Biological Series: Vol. 19 : No. 2 , Article 1. Available at: https://scholarsarchive.byu.edu/byuscib/vol19/iss2/1 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Brigham Young University Science Bulletin, Biological Series by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. S'/U/?- FJrpya Brigham Young University Science Bulletin *'''^ubrTry°°'" MAY271P74 HARVARD UNIVERSITAXONOMY OF THE GENUS CHLORIS (GRAMINEAE) by Dennis E. Anderson BIOLOGICAL SERIES — VOLUME XIX, NUMBER 2 MARCH 1974 /ISSN 0068-1024 BRIGHAM YOUNG UNIVERSITY SCIENCE BULLETIN BIOLOGICAL SERIES Editor: Stanley L. Welsh, Department of Botany, Brigham Young University, Provo, Utah Acting Editor: Vernon J. Tipton, Zoology Members of the Editorial Board: Ferron L. Andersen, Zoology Joseph R. Murdock, Botany WiLMER W. Tanner, Zoology Ex officio Members: A. Lester Allen, Dean, College of Biological and Agricultural Sciences Ernest L. Olson, Director, Brigham Young University Press The Brigham Young University Science Bulletin, Biological Series, publishes acceptable papers, particularly large manuscripts, on all phases of biology. Separate numbers and back volumes can be purchased from University Press Marketing, Brigham Young University, Provo, Utah 84602. -

Karyotype Evolution in Eleusininae Dumort. (Poaceae, Chloridoideae, Cynodonteae): a Phylogenetic and Cytogenetic Approach Focusing on the Genus Cynodon Rich
1 RAQUEL BEZERRA CHIAVEGATTO Karyotype evolution in Eleusininae Dumort. (Poaceae, Chloridoideae, Cynodonteae): a phylogenetic and cytogenetic approach focusing on the genus Cynodon Rich. LAVRAS-MG 2018 2 RAQUEL BEZERRA CHIAVEGATTO Karyotype evolution in Eleusininae Dumort. (Poaceae, Chloridoideae, Cynodonteae): a phylogenetic and cytogenetic approach focusing on the genus Cynodon Rich. Tese apresentada à Universidade Federal de Lavras, como parte das exigências do Programa de Pós Graduação em Botânica Aplicada, área de concentração em Botânica Aplicada, para a obtenção do título de Doutor. Profa. Dra. Vânia Helena Techio - DBI/UFLA Orientadora Prof. Dr. Lorenzo Peruzzi Coorientador Dr. Flávio Rodrigo Gandolfi Benites - Embrapa Gado de Leite Coorientador LAVRAS-MG 2018 3 Ficha catalográfica elaborada pelo Sistema de Geração de Ficha Catalográfica da Biblioteca Universitária da UFLA, com dados informados pelo(a) próprio(a) autor(a). Chiavegatto, Raquel Bezerra. Karyotype evolution in Eleusininae Dumort. (Poaceae, Chloridoideae, Cynodonteae): a phylogenetic and cytogenetic approach focusing on the genus Cynodon Rich. / Raquel Bezerra Chiavegatto. - 2018. 68 p. : il. Orientador(a): Vânia Helena Techio. Coorientador(a): Lorenzo Peruzzi, Flávio Rodrigo Gandolfi Benites. Tese (doutorado) - Universidade Federal de Lavras, 2018. Bibliografia. 1. Gramíneas forrageiras. 2. Evolução cromossômica. 3. Reconstrução do estado ancestral. I. Techio, Vânia Helena. II. Peruzzi, Lorenzo. III. Benites, Flávio Rodrigo Gandolfi. IV. Título. O conteúdo desta obra é de responsabilidade do(a) autor(a) e de seu orientador(a). 4 RAQUEL BEZERRA CHIAVEGATTO Karyotype evolution in Eleusininae Dumort. (Poaceae, Chloridoideae, Cynodonteae): a phylogenetic and cytogenetic approach focusing on the genus Cynodon Rich. Tese apresentada à Universidade Federal de Lavras, como parte das exigências do Programa de Pós Graduação em Botânica Aplicada, área de concentração em Botânica Aplicada, para a obtenção do título de Doutor. -
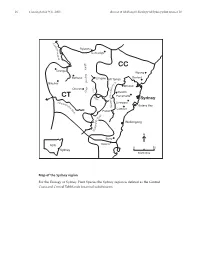
Part 10 ESP Intro
16 Cunninghamia 9(1): 2005 Benson & McDougall, Ecology of Sydney plant species 10 M a c q u Rylstone a r i e Coricudgy R i v e r e g n CC a Orange R Wyong g n i Gosford Bathurst d i Lithgow v Mt Tomah i Blayney D R. y r Windsor C t u a o b Oberon s e x r e s G k Penrith w a R Parramatta CT H i ve – Sydney r n a Abe e Liverpool rcro p m e b Botany Bay ie N R Camden iv Picton er er iv R y l l i Wollongong d n o l l o W N Berry NSW Nowra 050 Sydney kilometres Map of the Sydney region For the Ecology of Sydney Plant Species the Sydney region is defined as the Central Coast and Central Tablelands botanical subdivisions. Cunninghamia 9(1): 2005 Benson & McDougall, Ecology of Sydney plant species 10 17 Ecology of Sydney plant species Part 10 Monocotyledon families Lemnaceae to Zosteraceae Doug Benson and Lyn McDougall Royal Botanic Gardens and Domain Trust, Sydney, AUSTRALIA 2000. Email: [email protected] Abstract: Ecological data in tabular form are provided on 668 plant species of the families Lemnaceae to Zosteraceae, 505 native and 163 exotics, occurring in the Sydney region, defined by the Central Coast and Central Tablelands botanical subdivisions of New South Wales (approximately bounded by Lake Macquarie, Orange, Crookwell and Nowra). Relevant Local Government Areas are Auburn, Ashfield, Bankstown, Bathurst, Baulkham Hills, Blacktown, Blayney, Blue Mountains, Botany, Burwood, Cabonne, Camden, Campbelltown, Canada Bay, Canterbury, Cessnock, Crookwell, Evans, Fairfield, Greater Lithgow, Gosford, Hawkesbury, Holroyd, Hornsby, Hunters Hill, Hurstville, Kiama, Kogarah, Ku-ring-gai, Lake Macquarie, Lane Cove, Leichhardt, Liverpool, Manly, Marrickville, Mosman, Mulwaree, North Sydney, Oberon, Orange, Parramatta, Penrith, Pittwater, Randwick, Rockdale, Ryde, Rylstone, Shellharbour, Shoalhaven, Singleton, South Sydney, Strathfield, Sutherland, Sydney City, Warringah, Waverley, Willoughby, Wingecarribee, Wollondilly, Wollongong, Woollahra and Wyong. -

Chloridoideae- Poaceae)
UNIVERSIDAD NACIONAL DEL LITORAL Facultad de Bioquímica y Ciencias Biológicas Tesis para la obtención del Grado Académico de Doctor en Ciencias Biológicas EVOLUCIÓN, DESARROLLO Y ESTRUCTURA DE LAS INFLORESCENCIAS EN LA SUBTRIBU ELEUSININAE (CHLORIDOIDEAE- POACEAE) Sebastián E. Muchut Director de Tesis: Abelardo C. Vegetti Co-director de Tesis: Renata Reinheimer Cátedra de Morfología Vegetal Departamento de Biología Vegetal Facultad de Ciencias Agrarias – Universidad Nacional del Litoral -2017- Agradecimientos Quiero expresar mi agradecimiento a la Facultad de Bioquímica y Ciencias Biológicas de la Universidad Nacional del Litoral (UNL) por permitirme realizar el “Doctorado en Ciencias Biológicas”, especialmente al personal de la oficina de posgrado por resolver siempre atentamente mis inquietudes. A la Facultad de Ciencias Agrarias (FCA-UNL) por brindarme todas las instalaciones y herramientas necesarias para la realización de esta tesis. A la Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) y al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), por haberme otorgado las becas de posgrado que me permitieron dedicarme exclusivamente a la tesis doctoral. A mis directores de tesis, los Dres. Abelardo Vegetti y Renata Reinheimer por darme la oportunidad de iniciarme en la investigación científica, por enseñarme todo lo referido a la morfología, taxonomía, filogenia y evolución de gramíneas, por sus instrucciones con respecto a la escritura científica, por sus consejos, paciencia, y por todo el apoyo brindado durante estos años. A Abelardo, por haberme dado la posibilidad de integrar su grupo de trabajo, transmitirme su pasión por la investigación, aconsejarme constantemente e instruirme sobre el mundo científico. A la cátedra de Morfología Vegetal, por integrarme al grupo desde el primer día y hacerme sentir uno más de ellos, por los momentos académicos y recreativos compartidos. -

Analysis, Concerns, and Challenges
ASSESSMENT OF USER NEEDS OF PRIMARY BIODIVERSITY DATA: ANALYSIS, CONCERNS, AND CHALLENGES Arturo H. Ariño, Vishwas Chavan, Daniel P. Faith ANNEX: FREE-TEXT ANSWERS TO THE CNA-TG SURVEY Arturo H. Ariño1*, Burke Chih-Jen Ko2 (1) Department of Zoology and Ecology, University of Navarra, E-31080 Pamplona, Spain; [email protected] (2) Biodiversity Research Center, Academia Sinica, 128 Sec. 2, Academia Rd., Nangang Dist., Taipei City 115, Taiwan (R.O.C.); [email protected] * Corresponding author SUMMARY This is a comprehensive list of all answers collected from the CNA-TG survey which were allowed in free-text form, i.e. not pre-coded as multiple choice, tick-boxes or radio buttons. These have been arranged by question, then alphabetically. All six language-specific surveys have been merged together. The small number and letter code at the right of each answer corresponds to the respondent’s automatic ID and language of the original survey. NOTES 1. Answers in italics coming from the original Chinese, Spanish, French and Russian versions are translations into English by Burke Chih-Jen Ko (CN) and Arturo H. Ariño (ES, FR, RU). 2. Some discrete answers (i.e. "yes", "OK", tick, etc.) have been recoded into homogeneous bits. 3. Some typos or obvious mistakes have been summarily corrected for analysis and may not reflect the verbatim answer as recorded. 4. Some references have been reformatted for homogeneity. 5. As a rule, full literature references have not been translated from the ES, FR and RU versions except where noted (translated titles appearing in brackets by the original version). -

Identifying Western Australian Summer Weeds
IDENTIFYING WESTERN AUSTRALIAN SUMMER WEEDS MANUAL WESTERN Title: Identifying Western Australian Summer Weeds Manual GRDC Project Code: 9176937 – Early and Effective Summer Weed Control Author: Andrew Storrie, Agronomo © Grains Research and Development Corporation All material published in this publication is copyright protected and may not be reproduced in any form without written permission from GRDC. Published January 2019. ISBN: 978-1-921779-73-2 (online); 978-1-921779-7409 (print) Copies of this report can be downloaded from www.grdc.com.au or a hard copy can be obtained from: GRDC Western Region Office Suite 5, 2a Brodie Hall Drive BENTLEY WA 6102 Ph: 08 9230 4600; email: [email protected] GRDC contact details: Ms Maureen Cribb Integrated Publications Manager PO Box 5367 KINGSTON ACT 2604 Email: [email protected] Design and production: Coretext, www.coretext.com.au Front cover image: Richard Holzknecht Disclaimer: Any recommendations, suggestions or opinions contained in this publication do not necessarily represent the policy or views of the Grains Research and Development Corporation. No person should act on the basis of the contents of this publication without first obtaining specific, independent professional advice. The Grains Research and Development Corporation may identify products by proprietary or trade names to help readers identify particular types of products. We do not endorse or recommend the products of any manufacturer referred to. Other products may perform as well as or better than those specifically referred to. GRDC will not be liable for any loss, damage, cost or expense incurred or arising by reason of any person using or relying on the information in this manual. -

Fol Ia Botanica Extremadurensis
Volumen 13(2) OCTUBRE 2019 Guia excursão geobotânica FOLIA BOTANICA EXTREMADURENSIS FOLIA XIII Seminário Internacional Gestão Coordinación: Francisco Mª Vázquez Pardo Secretaría: Francisco Márquez García Equipo de edición: David García Alonso, María José Guerra Barrena y Francisco Márquez García Equipo de redacción y revisión de textos: José Blanco Salas David García Alonso Francisco Márquez García José Luis Pérez Chiscano Carlos Pinto Gomes Francisco M. Vázquez Pardo Ilustración de portada: Espiguillas de Alopecurus arundinaceus subsp. arundinaceus Poiret Edita: Centro de Investigaciones Científicas y Tecnológicas de Extremadura (CICYTEX). ISSN: 1887-6587 Depósito legal: BA-178-07 Diseño: Grupo HABITAT. Imprime: IBERPRINT, Montijo (Badajoz, España). Unidad de Biodiversidad Vegetal. Herbario HSS. Instituto de Investigaciones Agrarias “Finca La Orden”. A-V, km 372. 06187 GUADAJIRA (BADAJOZ (España)). Centro de Investigaciones Científicas y Tecnológicas de Extremadura (CICYTEX) Consejería de Economía, Ciencia y Agenda Digital. Junta de Extremadura. FOLIA BOTANICA EXTREMADURENSIS Vol. 13(2) OCTUBRE 2019 Coordinación: Francisco Mª Vázquez Pardo Secretaría: Francisco Márquez García Equipo de edición: David García Alonso, Francisco Márquez García, y María José Guerra Barrena. Equipo de redacción y revisión de textos: José Blanco Salas David García Alonso Francisco Márquez García José Luis Pérez Chiscano Carlos Pinto Gomes Francisco Mª Vázquez Pardo Ilustración de portada: Espiguillas de Alopecurus arundinaceus Poiret Edita: Centro de Investigaciones Científicas y Tecnológicas de Extremadura (CICYTEX). ISSN: 1887-6587 Depósito legal: BA-178-07 Diseño: Grupo HABITAT. Imprime: IBERPRINT. Montijo (Badajoz, España). Unidad de Biodiversidad Vegetal. Herbario HSS. Instituto de Investigaciónes Agrarias “La Orden”. A-V, km 372. 06187 GUADAJIRA (BADAJOZ (España)). Centro de Investigaciones Científicas y Tecnológicas de Extremadura (CICYTEX). -

El Género Chloris SW (Poaceae: Chloridoideae)
Acta Botanica Mexicana 112: 95-147 (2015) EL GÉNERO CHLORIS SW. (POACEAE: CHLORIDOIDEAE) EN MÉXICO ROSA CErrOS-TLATILPA1, 4, MARÍA ELENA SIQUEIROS DELGADO2 Y ELIZABETH M. SKENDZIC3 1Universidad Autónoma del Estado de Morelos, Facultad de Ciencias Biológicas, Avenida Universidad 1001, Colonia Chamilpa, 62210 Cuernavaca, Morelos, México. 2Universidad Autónoma de Aguascalientes, Departamento de Biología, Centro Básico, Avenida Universidad 940, 20000 Aguascalientes, Aguascalientes, México. 3Kutztown University of Pennsylvania, Biology Department, 15200 Kutztown Road, Kutztown, Pennsylvania 19530, USA. 4Autor para la correspondencia: [email protected] RESUMEN En este estudio se reconocen 12 especies de Chloris para el territorio de la República. Cinco son cosmopolitas, cinco americanas y dos de ellas se restringen a México y E.U.A., una está en América y en Australia, mientras que C. truncata se reporta por primera vez de nuestro país. Chloris barbata, C. gayana y C. virgata son consideradas invasoras para el país. Se incluye una clave para su identificación, descripciones y mapas de distribución. El género ocupa una variedad de hábitats, desde zonas con disturbio hasta áreas conservadas. Palabras clave: especies invasoras, diversidad, florística, nuevos registros, revisión. ABSTRACT Twelve species of Chloris from Mexico are recognized in this study. Five are cosmopolitan species, five are American, two are restricted to Mexico and USA, one occurs in America and Australia, whereas C. truncata is recorded for the first time in Mexico. Chloris barbata, C. gayana, and C. virgata are considered invasive species in the country. An identification key, descriptions and distribution maps are included. The genus occupies a variety of habitats, growing from disturbed zones to conserved areas.