Development of the Red Maple Understory in Northeastern Oak
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Genetic and Phenotypic Characterization of Figured Wood in Poplar
Genetic and Phenotypic Characterization of Figured Wood in Poplar Youran Fan1,2, Keith Woeste1,2, Daniel Cassens1, Charles Michler1,2, Daniel Szymanski3, and Richard Meilan1,2 1Department of Forestry and Natural Resources, 2Hardwood Tree Improvement and Regeneration Center, and 3Department of Agronomy; Purdue University, West Lafayette, Indiana 47907 Abstract Materials and Methods When “Curly Aspen” (Populus canescens) was first Preliminary Results characterized in the early 1940’s[1], it attracted the attention from the wood-products industry because Genetically engineer commercially 1) Histological sections reveal that “Curly Aspen” has strong “Curly Aspen” produces an attractive veneer as a important trees to form figure. ray flecks (Fig. 10) but this is not likely to be responsible result of its figured wood. Birdseye, fiddleback and for the figure seen. quilt are other examples of figured wood that are 2) Of the 15 SSR primer pairs[6, 7, 8] tested, three have been commercially important[2]. These unusual grain shown to be polymorphic. Others are now being tested. patterns result from changes in cell orientation in Figure 6. Pollen collection. Branches of Figure 7. Pollination. Branches Ultimately, our genetic fingerprinting technique will allow “Curly Aspen” were “forced” to shed collected from a female P. alba us to distinguish “Curly Aspen” from other genotypes. the xylem. Although 50 years have passed since Figure 1. Birdseye in maple. pollen under controlled conditions. growing at Iowa State University’s finding “Curly Aspen”, there is still some question Rotary cut, three-piece book McNay Farm (south of Lucas, IA). 3) 17 jars of female P. alba branches have been pollinated match (origin: North America). -

The Story About Understory Why Everyone on Bald Head Island Should Care
Bald Head Association 910-457-4676 • www.BaldHeadAssociation.com 111 Lighthouse Wynd • PO Box 3030, Bald Head Island, NC 28461 The Story About Understory Why Everyone on Bald Head Island Should Care Bald Head Island is truly unique. As a barrier island, it at ground level. Bald Head Island’s latitudinal position is bordered by the Atlantic Ocean and Cape Fear River. BHI has supports both northern and southern species of plants.” 14 miles of pristine beaches; 12,400 acres, of which 10,000 acres Important understory plants include vines, small plants and are protected; over 244 species of birds, including the bald eagle; trees, mosses, lichens and even weeds. All are needed to have and the North Carolina Forest Preserve of nearly 200 acres. a healthy forest habitat. Many extreme forces of nature that have helped shape this The BHIC explains, “Vines play an important role. The very special island include hurricane-force winds, salt water and vines and herbaceous plants intertwine, further developing the salt spray, flooding and drought. There is another force of nature structural integrity of the forest and forming pockets of vegetation that impacts BHI — humans — and with us, development. that provide a base for songbirds to build nests. The vines twist The Bald Head Association is mandated by its Covenants to around the canopy and are the secret to wind protection. These help sustain BHI by managing the buildout of homes and by vines actually weave together the canopy so blowing winds don’t managing vegetation trimming/removal, to help protect its penetrate through the top layer and keep homes, plants, and members’ property values. -

Old-Growth Forests
Pacific Northwest Research Station NEW FINDINGS ABOUT OLD-GROWTH FORESTS I N S U M M A R Y ot all forests with old trees are scientifically defined for many centuries. Today’s old-growth forests developed as old growth. Among those that are, the variations along multiple pathways with many low-severity and some Nare so striking that multiple definitions of old-growth high-severity disturbances along the way. And, scientists forests are needed, even when the discussion is restricted to are learning, the journey matters—old-growth ecosystems Pacific coast old-growth forests from southwestern Oregon contribute to ecological diversity through every stage of to southwestern British Columbia. forest development. Heterogeneity in the pathways to old- growth forests accounts for many of the differences among Scientists understand the basic structural features of old- old-growth forests. growth forests and have learned much about habitat use of forests by spotted owls and other species. Less known, Complexity does not mean chaos or a lack of pattern. Sci- however, are the character and development of the live and entists from the Pacific Northwest (PNW) Research Station, dead trees and other plants. We are learning much about along with scientists and students from universities, see the structural complexity of these forests and how it leads to some common elements and themes in the many pathways. ecological complexity—which makes possible their famous The new findings suggest we may need to change our strat- biodiversity. For example, we are gaining new insights into egies for conserving and restoring old-growth ecosystems. canopy complexity in old-growth forests. -

Comparison of Oak and Sugar Maple Distribution and Regeneration in Central Illinois Upland Oak Forests
COmparisON OF OaK AND Sugar MAPLE DistriBUTION AND REGENEratiON IN CEntral ILLINOIS UPLAND OaK FOREsts Peter J. Frey and Scott J. Meiners1 Abstract.—Changes in disturbance frequencies, habitat fragmentation, and other biotic pressures are allowing sugar maple (Acer saccharum) to displace oak (Quercus spp.) in the upland forest understory. The displacement of oaks by sugar maples represents a major management concern throughout the region. We collected seedling microhabitat data from five upland oak forest sites in central Illinois, each differing in age class or silvicultural treatment to determine whether oaks and maples differed in their microhabitat responses to environmental changes. Maples were overall more prevalent in mesic slope and aspect positions. Oaks were associated with lower stand basal area. Both oaks and maples showed significant habitat partitioning, and environmental relationships were consistent across sites. Results suggest that management intensity for oak in upland forests could be based on landscape position. Maple expansion may be reduced by concentrating mechanical treatments in expected areas of maple colonization, while using prescribed fire throughout stands to promote oak regeneration. INTRODUCTION Historically, white oak (Quercus alba) dominated much of the midwestern and eastern U.S. hardwood forests (Abrams and Nowacki 1992, Franklin et al. 1993). Oak is classified as an early successional forest species, and many researchers agree that oak populations were maintained by Native American or lightning-initiated fires (Abrams 2003, Abrams and Nowacki 1992, Hutchinson et al. 2008, Moser et al. 2006, Nowacki and Abrams 2008, Ruffner and Groninger 2006, Shumway et al. 2001). These periodic low to moderate surface fires favored the ecophysiological attributes of oak over those of fire-sensitive, shade-tolerant tree species, thereby continually resetting succession and allowing oaks and other shade-intolerant species to persist in both the canopy and understory (Abrams 2003, Abrams and Nowacki 1992, Crow 1988, Franklin et al. -

Sugar Maple - Oak - Hickory Forest State Rank: S3 - Vulnerable
Sugar Maple - Oak - Hickory Forest State Rank: S3 - Vulnerable Mesic Forest (RMF): Sugar Maple - Oak - Hickory Forests are most occurrences of RMF diverse forests in central and eastern in Massachusetts are west Massachusetts where conditions, of the Connecticut River including nutrient richness, support Valley. The presence of Northern Hardwood species mixed with multiple species of species of Oak - Hickory Forests; hickories and oaks in SMOH is a main The herbaceous layer varies from sparse difference between these to intermittent, with sparse spring two types. Broad-leaved ephemerals that may include bloodroot or Woodland-sedge is close trout-lily. Later occurring species may to being an indicator of include wild geranium, herb Robert, wild SMOH. RMF is Rock outcrops in the spring in Sugar Maple - licorice, maidenhair fern, bottlebrush Oak - Hickory Forest area. Photo: Patricia characterized by very Swain, NHESP. grass, and white wood aster. Broad- dense herbaceous growth of spring leaved, semi-evergreen broad-leaved ephemerals; SMOH shares some of the Description: Sugar Maple - Oak - woodland-sedge is close to an indicator of species but with fewer individuals of Hickory Forests occur in or east of the the community. Witch hazel, hepaticas, fewer species. SMOH has evergreen Connecticut River Valley in and wild oats usually occur in transitions ferns, Christmas fern and wood ferns, that Massachusetts. They are associated with to surrounding forest types. RMF lack. Oak - Hickory Forests and outcrops of circumneutral rock and slopes Dry, Rich Oak Forests/Woodlands lack below them that have more nutrients than abundant sugar maple, basswood, and are available in the surrounding forest. -

Understory Forest Monitoring: a Guide for Small Forest Managers
Understory Forest Monitoring: A Guide for Small Forest Managers With funding provided by BEFORE: A dense monoculture of invasive periwinkle (Vinca species). AFTER: A diverse mix of native vegetation beginning to establish. It will mature into a lush forest Á oor of native wildÁ owers. Managing a healthy forest is not just about having healthy trees. Foresters, scientists, and private woodland owners have an interest in what is growing on the forest Á oor. A healthy understory can offer: • Flowers for native pollinators • Food for wildlife • Resiliency against invasion by forest weeds • Organic material to build healthier soil • Stable soil that doesn’t erode into nearby streams • Beautiful views while recreating in the forest Just like taking inventory of the forest, measuring tree diameters and spacing, it is also important to monitor the changes over time on the forest Á oor. Herbaceous forest plants tend to reach their peak bloom and cover in mid to late spring (mid-May to mid-June in western Oregon). It’s best to monitor forest understory consistently at this time of year. There are dozens of ways to monitor the understory plants in a forest. Two effective methods are described here. Point intercept method This method is performed by laying out a straight line transect in the forest and documenting what is growing on the forest Á oor at one- foot increments along the transect. West Multnomah Soil & Water Conservation District often sets up multiple transects that are 33’ in length giving us 33 data points per transect. The data gathered from the plots can be used to À nd the average cover of different vegetation types in your forest. -

Accelerating the Development of Old-Growth Characteristics in Second-Growth Northern Hardwoods
United States Department of Agriculture Accelerating the Development of Old-growth Characteristics in Second-growth Northern Hardwoods Karin S. Fassnacht, Dustin R. Bronson, Brian J. Palik, Anthony W. D’Amato, Craig G. Lorimer, Karl J. Martin Forest Northern General Technical Service Research Station Report NRS-144 February 2015 Abstract Active management techniques that emulate natural forest disturbance and stand development processes have the potential to enhance species diversity, structural complexity, and spatial heterogeneity in managed forests, helping to meet goals related to biodiversity, ecosystem health, and forest resilience in the face of uncertain future conditions. There are a number of steps to complete before, during, and after deciding to use active management for this purpose. These steps include specifying objectives and identifying initial targets, recognizing and addressing contemporary stressors that may hinder the ability to meet those objectives and targets, conducting a pretreatment evaluation, developing and implementing treatments, and evaluating treatments for success of implementation and for effectiveness after application. In this report we discuss these steps as they may be applied to second-growth northern hardwood forests in the northern Lake States region, using our experience with the ongoing managed old-growth silvicultural study (MOSS) as an example. We provide additional examples from other applicable studies across the region. Quality Assurance This publication conforms to the Northern Research Station’s Quality Assurance Implementation Plan which requires technical and policy review for all scientific publications produced or funded by the Station. The process included a blind technical review by at least two reviewers, who were selected by the Assistant Director for Research and unknown to the author. -
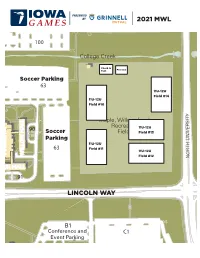
2021 Mwl 2 L
Schilletter - University SCHILLETTER - UNIVERSITY VILLAGE201R COMMUNITY 201B 201D 201N CENTER D A O APPLIED R GOLF COURSE SCIENCE 201P S MAINTENANCE T COMPLEX IV T OFFICE O T LONG ROAD S G DR NBUR IVE 201M BLANK201CE 201K 104 CLUBHOUSE APPLIED D SCIENCE A COMPLEX I O R 201L 102 G 201C APPLIED 103 N O SCIENCE L COMPLEX III APPLIED D SCIENCE 201J A O COMPLEX II 201H R WASTE S T CHEMICAL T 201F D 201G O HANDLING T 101 A S SENSITIVE BUILDING Veenker Memorial Golf Course O INSTRUMENT R FACILITY E LYNN G FUHRER N A LODGE T S 205 201E 201D 201B ISU FAMILY 200 RESOURCE CENTER 205 RIVE 201A BRUNER D Iow ay C reek D A O R L L O H Io C w S ay Cre ek Disc Golf Course 112 L 13 Veenker Memorial Golf Course TH S TRE Furnam Aquatic Center ET EXTENSION 4-H (City Of Ames) T 125 REE YOUTH BUILDING H ST 13T 112 J 124A 112 A 112 J 112 H EH&S ONTARIO STREET 122A ONTARIO STREET SERVICES 120A ROAD STANGE BUILDING 124 FRED115ERIKSEN 112 K 121 BLDG SERVICES COURT ADMINISTRATIVE 112 G HAWTHORN COURT COMMUNITY FREDERIKSEN WANDA DALEY DRIVE DRIVE CENTER COURT 112 K 112 K 122 LIBRARY 120 DOE STORAGE WAREHOUSE FACILITY 119 DOE DOE CONST D 112 D-1 MECH A DOE O R 29 MAINT DOE R PRINTING & E SHOP B ROY J. CARVER PUBLICATIONS31 A CO-LABORATORY BUILDING H 112N 29 NORTH CHILLED 112 B 112 F WATER PLANT ear Creek 112 C 112 D-2 Cl 28 28 Union Pacific Railroad 35 30 27 28A 33 Pammel Woods 12 MOLECULAR 28A FIRE SERVICE BIOLOGY BUILDING COMMUNICATIONS 32 A BUILDING METALS TRANSPORTATION S ADVANCED BUILDING 28A 110 DEVELOPMENT C RUMINANT I 32 TEACHING & HORSE BARN BUILDING T NUTRITION SERVICES E RESEARCH 79 STANGE ROAD STANGE N LAB Cemetery E BUILDING WINLOCK ROAD G LABORATORY MACH ADVANCED LAB SYSTEMS 11W 11E PAMMEL DRIVE NORTH UNIVERSITY BOULEVARD PAMMEL DRIVE SPEDDING WILHELM NATIONAL NATIONAL HYLAND AVENUE HYLAND 11 22 HACH HALL HALL HALL SCIENCE SWINE RSRCH LAB FOR AG. -

Folivory of Vine Maple in an Old-Growth Douglas-Fir-Western Hemlock Forest
3589 David M. Braun, Bi Runcheng, David C. Shaw, and Mark VanScoy, University of Washington, Wind River Canopy Crane Research Facility, 1262 Hemlock Rd., Carson, Washington 98610 Folivory of Vine Maple in an Old-growth Douglas-fir-Western Hemlock Forest Abstract Folivory of vine maple was documented in an old-growth Douglas-fir-western hemlock forest in southwest Washington. Leaf consumption by lepidopteran larvae was estimated with a sample of 450 tagged leaves visited weekly from 7 May to 11 October, the period from bud break to leaf drop. Lepidopteran taxa were identified by handpicking larvae from additional shrubs and rearing to adult. Weekly folivory peaked in May at 1.2%, after which it was 0.2% to 0.7% through mid October. Cumulative seasonal herbivory was 9.9% of leaf area. The lepidopteran folivore guild consisted of at least 22 taxa. Nearly all individuals were represented by eight taxa in the Geometridae, Tortricidae, and Gelechiidae. Few herbivores from other insect orders were ob- served, suggesting that the folivore guild of vine maple is dominated by these polyphagous lepidopterans. Vine maple folivory was a significant component of stand folivory, comparable to — 66% of the folivory of the three main overstory conifers. Because vine maple is a regionally widespread, often dominant understory shrub, it may be a significant influence on forest lepidopteran communities and leaf-based food webs. Introduction tract to defoliator outbreaks, less is known about endemic populations of defoliators and low-level Herbivory in forested ecosystems consists of the folivory. consumption of foliage, phloem, sap, and live woody tissue by animals. -

E666134Z Enhance Development of the Forest Understory to Create
CONSERVATION ENHANCEMENT ACTIVITY E666134Z Enhance development of the forest understory to create conditions resistant to pests Conservation Practice 666: Forest Stand Improvement APPLICABLE LAND USE: Forest RESOURCE CONCERN ADDRESSED: Degraded Plant Condition PRACTICE LIFE SPAN: 10 Years Enhancement Description: Forest stand improvement that manages the structure and composition of overstory and understory vegetation to reduce vulnerability to damage by insects and diseases of forest trees. Managing the understory vegetation will also reduce the risk of wildfire, and promote development of herbaceous plants that benefit wildlife. This enhancement provides for management of the understory vegetation in a forested area, using mechanical, chemical or manual methods to improve the plant species mix and the health of the residual vegetation. These practices encourage the development of a less dense forested ecosystem. The canopy gaps and open understory allow for air circulation that reduces the incidence of disease, and the improved health of the residual trees increases their ability to withstand insect attacks. Forest stand improvement (FSI) activities are used to remove trees of undesirable species, form, quality, condition, or growth rate. The quantity and quality of forest for wildlife and/or timber production will be increased by manipulating stand density and structure. These treatments can also reduce wildfire hazards, improve forest health, restore natural plant communities, and achieve or maintain a desired native understory plant community for soil health, wildlife, grazing, and/or browsing. Criteria: States will apply general criteria from the NRCS National Conservation Practice Standard (CPS) 666 as listed below, and additional criteria as required by the NRCS State Office. The enhancement will be applied to sites which have an uncharacteristically dense understory of shrubs and small trees that limit development of ground cover. -

4-H Wood Science Leader Guide Glossary of Woodworking Terms
4-H Wood Science Leader Guide Glossary of Woodworking Terms A. General Terms B. Terms Used in the Lumber Industry d—the abbreviation for “penny” in designating nail boards—Lumber less than 2 inches in nominal size; for example, 8d nails are 8 penny nails, 2½” long. thickness and 1 inch and wider in width. fiber—A general term used for any long, narrow cell of board foot—A measurement of wood. A piece of wood wood or bark, other than vessels. that is 1 foot long by 1 foot wide by 1 inch thick. It can also be other sizes that have the same total amount grain direction—The direction of the annual rings of wood. For example, a piece of wood 2 feet long, showing on the face and sides of a piece of lumber. 6 inches wide, and 1 inch thick; or a piece 1 foot long, hardwood—Wood from a broad leaved tree and 6 inches wide, and 2 inches thick would also be 1 board characterized by the presence of vessels. (Examples: foot. To get the number of board feet in a piece of oak, maple, ash, and birch.) lumber, measure your lumber and multiply Length (in feet) x Width (in feet) x Thickness (in inches). The heartwood—The older, harder, nonliving portion of formula is written: wood. It is usually darker, less permeable, and more durable than sapwood. T” x W’ x L’ T” x W’ x L’ = Board feet or = Board feet 12 kiln dried—Wood seasoned in a humidity and temperature controlled oven to minimize shrinkage T” x W’ x L’ or = Board feet and warping. -

Self-Guided Neighborhood Tree Tour Created by Bhopesh Bassi
BELLEVUE NEIGHBORHOOD TREE AMBASSADOR PROGRAM Self-Guided Neighborhood Tree Tour Created by Bhopesh Bassi Neighborhood: Kelsey Creek Park and Farm Starting point: Kelsey Creek Park and Farm parking lot at 410 130th Place SE Summary/Theme: Introduction to plant and animal ecosystem in and around Kelsey Creek Park and Farm. Kelsey Creek Farm Park in Bellevue is a largely overlooked gem. While the park is tucked in the heart of neighborhoods, it feels like an isolated country woods experience. You may have the impression of having traveled back in time as you move from the modern parking lot to a more rural feel as you hike into the forest. Kelsey Creek is Seattle's largest watershed. It runs right down the middle of the park, providing habitat for spawning Salmon, Peamouth and for an abundant population of migratory and resident birds. All lifeform here depends on and contributes to each other and the park is a rich ecosystem. Bring your binoculars along if you also want to watch birds on this tour. BELLEVUE NEIGHBORHOOD TREE AMBASSADOR PROGRAM This tree tour was developed by one of Bellevue’s Neighborhood Tree Ambassador volunteers. The goal of the Neighborhood Tree Ambassador program is to help build community support for trees in Bellevue. Trees are an important part of our community because they provide significant health and environmental benefits. Trees: • Remove pollutants from the air and water • Reduce stress and improve focus • Lower air temperature • Pull greenhouse gases from the atmosphere • Reduce flooding and erosion caused by rain Bellevue has a goal to achieve a 40% tree canopy across the entire city.