Factor-Induced Receptor Ubiquitination and Mediates
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Promoter Janus Kinase 3 Proximal Characterization and Analysis Of
The Journal of Immunology Characterization and Analysis of the Proximal Janus Kinase 3 Promoter1 Martin Aringer,2*† Sigrun R. Hofmann,2* David M. Frucht,* Min Chen,* Michael Centola,* Akio Morinobu,* Roberta Visconti,* Daniel L. Kastner,* Josef S. Smolen,† and John J. O’Shea3* Janus kinase 3 (Jak3) is a nonreceptor tyrosine kinase essential for signaling via cytokine receptors that comprise the common ␥-chain (␥c), i.e., the receptors for IL-2, IL-4, IL-7, IL-9, IL-15, and IL-21. Jak3 is preferentially expressed in hemopoietic cells and is up-regulated upon cell differentiation and activation. Despite the importance of Jak3 in lymphoid development and immune function, the mechanisms that govern its expression have not been defined. To gain insight into this issue, we set out to characterize the Jak3 promoter. The 5-untranslated region of the Jak3 gene is interrupted by a 3515-bp intron. Upstream of this intron and the transcription initiation site, we identified an ϳ1-kb segment that exhibited lymphoid-specific promoter activity and was responsive to TCR signals. Truncation of this fragment revealed that core promoter activity resided in a 267-bp fragment that contains putative Sp-1, AP-1, Ets, Stat, and other binding sites. Mutation of the AP-1 sites significantly diminished, whereas mutation of the Ets sites abolished, the inducibility of the promoter construct. Chromatin immunoprecipitation assays showed that histone acetylation correlates with mRNA expression and that Ets-1/2 binds this region. Thus, transcription factors that bind these sites, especially Ets family members, are likely to be important regulators of Jak3 expression. -

Calycosin Down-Regulates C-Met to Suppress Development of Glioblastomas
J Biosci (2019) 44:96 Ó Indian Academy of Sciences DOI: 10.1007/s12038-019-9904-4 (0123456789().,-volV)(0123456789().,-volV) Calycosin down-regulates c-Met to suppress development of glioblastomas , XIAOHU NIE ,YUE ZHOU* ,XIAOBING LI,JIE XU,XUYAN PAN,RUI YIN and BIN LU Department of Neurosurgery, Huzhou Central Hospital, Huzhou, Zhejiang, People’s Republic of China *Corresponding author (Email, [email protected]) These authors contributed equally to this work. MS received 15 October 2018; accepted 9 June 2019; published online 7 August 2019 The antitumor effect of calycosin has been widely studied, but the targets of calycosin against glioblastomas are still unclear. In this study we focused on revealing c-Met as a potential target of calycosin suppressing glioblastomas. In this study, suppressed-cell proliferation and cell invasion together with induced-cell apoptosis appeared in calycosin-treated U251 and U87 cells. Under treatment of calycosin, the mRNA expression levels of Dtk, c-Met, Lyn and PYK2 were observed in U87 cells. Meanwhile a western blot assay showed that c-Met together with matrix metalloproteinases-9 (MMP9) and phosphorylation of the serine/threonine kinase AKT (p-AKT) was significantly down-regulated by calycosin. Furthermore, overexpressed c-Met in U87 enhanced the expression level of MMP9 and p-AKT and also improved cell invasion. Additionally, the expression levels of c-Met, MMP9 and p-AKT were inhibited by calycosin in c-Met overex- pressed cells. However, an AKT inhibitor (LY294002) only effected on MMP9 and p-AKT, not on c-Met. These data collectively indicated that calycosin possibility targeting on c-Met and exert an anti-tumor role via MMP9 and AKT. -

Inhibition of DDR1-BCR Signalling by Nilotinib As a New Therapeutic
Inhibition of DDR1-BCR signalling by nilotinib as a new therapeutic strategy for metastatic colorectal cancer Maya Jeitany, Cédric Leroy, Priscillia Tosti, Marie Lafitte, Jordy Le Guet, Valérie Simon, Debora Bonenfant, Bruno Robert, Fanny Grillet, Caroline Mollevi, et al. To cite this version: Maya Jeitany, Cédric Leroy, Priscillia Tosti, Marie Lafitte, Jordy Le Guet, et al.. Inhibition of DDR1- BCR signalling by nilotinib as a new therapeutic strategy for metastatic colorectal cancer. EMBO Molecular Medicine, Wiley Open Access, 2018, 10 (4), pp.e7918. 10.15252/emmm.201707918. hal- 01872978 HAL Id: hal-01872978 https://hal.archives-ouvertes.fr/hal-01872978 Submitted on 12 Jan 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution| 4.0 International License Research Article Inhibition of DDR1-BCR signalling by nilotinib as a new therapeutic strategy for metastatic colorectal cancer Maya Jeitany1,†, Cédric Leroy1,2,3,†, Priscillia Tosti1,†, Marie Lafitte1, Jordy Le Guet1, Valérie Simon1, Debora Bonenfant2, Bruno Robert4, Fanny Grillet5, Caroline Mollevi4, Safia El Messaoudi4, Amaëlle Otandault4, Lucile Canterel-Thouennon4, Muriel Busson4, Alain R Thierry4, Pierre Martineau4, Julie Pannequin5, Serge Roche1,*,† & Audrey Sirvent1,†,** Abstract The current clinical management involves surgical removal of the primary tumour, often associated with chemotherapy. -

Biomarker Testing in Non- Small Cell Lung Cancer (NSCLC)
The biopharma business of Merck KGaA, Darmstadt, Germany operates as EMD Serono in the U.S. and Canada. Biomarker testing in non- small cell lung cancer (NSCLC) Copyright © 2020 EMD Serono, Inc. All rights reserved. US/TEP/1119/0018(1) Lung cancer in the US: Incidence, mortality, and survival Lung cancer is the second most common cancer diagnosed annually and the leading cause of mortality in the US.2 228,820 20.5% 57% Estimated newly 5-year Advanced or 1 survival rate1 metastatic at diagnosed cases in 2020 diagnosis1 5.8% 5-year relative 80-85% 2 135,720 survival with NSCLC distant disease1 Estimated deaths in 20201 2 NSCLC, non-small cell lung cancer; US, United States. 1. National Institutes of Health (NIH), National Cancer Institute. Cancer Stat Facts: Lung and Bronchus Cancer website. www.seer.cancer.gov/statfacts/html/lungb.html. Accessed May 20, 2020. 2. American Cancer Society. What is Lung Cancer? website. https://www.cancer.org/cancer/non-small-cell-lung-cancer/about/what-is-non-small-cell-lung-cancer.html. Accessed May 20, 2020. NSCLC is both histologically and genetically diverse 1-3 NSCLC distribution by histology Prevalence of genetic alterations in NSCLC4 PTEN 10% DDR2 3% OTHER 25% PIK3CA 12% LARGE CELL CARCINOMA 10% FGFR1 20% SQUAMOUS CELL CARCINOMA 25% Oncogenic drivers in adenocarcinoma Other or ADENOCARCINOMA HER2 1.9% 40% KRAS 25.5% wild type RET 0.7% 55% NTRK1 1.7% ROS1 1.7% Oncogenic drivers in 0% 20% 40% 60% RIT1 2.2% squamous cell carcinoma Adenocarcinoma DDR2 2.9% Squamous cell carcinoma NRG1 3.2% Large cell carcinoma -
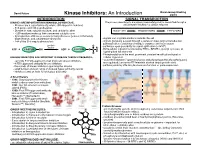
Kinase Inhibitors: an Introduction
David Peters Baran Group Meeting Kinase Inhibitors: An Introduction 2/2/19 INTRODUCTION SIGNAL TRANSDUCTION KINASES ARE IMPORTANT IN HUMAN BIOLOGY/DISEASE: The process describing how a signal (chemical/physical) is transmitted through a - Kinases are a superfamily of proteins (5th largest in humans) cell ultimately resulting in a cellular response - 518 genes and 106 pseudogenes - Diverse in size, subunit structure, and cellular location RECEPTOR TRANSDUCERS EFFECTORS - ~260 residues make up their conserved catalytic core - dysregulation of kinases occurs in many diseases (cancer, inflamatory, degenerative, and autoimmune diseases) - signals can originate inside or outside the cell - 244 of the 518 map to disease loci - signals generally passed through a series of steps (signal transduction pathway) often consisting of multiple enzymes and messengers protein O - pathways open possibility for signal aplification (>1x106) kinase ATP + PROTEIN OH ADP + PROTEIN O P O - Extracellular signals transduced by RTKs, GPCR’s, guanlyl cyclases, or ligand-gated Ion channels O - Phosphorylation is the most prominent covalent modification/signal in KINASES INHIBITORS ARE IMPORTANT IN DISEASE THERAPY/RESEARCH: cellular regulation - currently 51 FDA approved small molecule kinase inhibitors - “converter enzymes” (protein kinases and phosphoprotein phosphorlyases) - 4 FDA approved antibody kinase inhibitors are regulated; conserve ATP/maintain desired target protein state - thousands of known inhibitors spanning the kinome - pathway ends by affecting biomolecule -

Inhibition of Src Family Kinases and Receptor Tyrosine Kinases by Dasatinib: Possible Combinations in Solid Tumors
Published OnlineFirst June 13, 2011; DOI: 10.1158/1078-0432.CCR-10-2616 Clinical Cancer Molecular Pathways Research Inhibition of Src Family Kinases and Receptor Tyrosine Kinases by Dasatinib: Possible Combinations in Solid Tumors Juan Carlos Montero1, Samuel Seoane1, Alberto Ocaña2,3, and Atanasio Pandiella1 Abstract Dasatinib is a small molecule tyrosine kinase inhibitor that targets a wide variety of tyrosine kinases implicated in the pathophysiology of several neoplasias. Among the most sensitive dasatinib targets are ABL, the SRC family kinases (SRC, LCK, HCK, FYN, YES, FGR, BLK, LYN, and FRK), and the receptor tyrosine kinases c-KIT, platelet-derived growth factor receptor (PDGFR) a and b, discoidin domain receptor 1 (DDR1), c-FMS, and ephrin receptors. Dasatinib inhibits cell duplication, migration, and invasion, and it triggers apoptosis of tumoral cells. As a consequence, dasatinib reduces tumoral mass and decreases the metastatic dissemination of tumoral cells. Dasatinib also acts on the tumoral microenvironment, which is particularly important in the bone, where dasatinib inhibits osteoclastic activity and favors osteogenesis, exerting a bone-protecting effect. Several preclinical studies have shown that dasatinib potentiates the antitumoral action of various drugs used in the oncology clinic, paving the way for the initiation of clinical trials of dasatinib in combination with standard-of-care treatments for the therapy of various neoplasias. Trials using combinations of dasatinib with ErbB/HER receptor antagonists are being explored in breast, head and neck, and colorectal cancers. In hormone receptor–positive breast cancer, trials using combina- tions of dasatinib with antihormonal therapies are ongoing. Dasatinib combinations with chemother- apeutic agents are also under development in prostate cancer (dasatinib plus docetaxel), melanoma (dasatinib plus dacarbazine), and colorectal cancer (dasatinib plus oxaliplatin plus capecitabine). -

Receptor Signaling and Syk in the Initiation of B-Cell Antigen
Nonredundant Roles of Src-Family Kinases and Syk in the Initiation of B-Cell Antigen Receptor Signaling This information is current as Ondrej Stepanek, Peter Draber, Ales Drobek, Vaclav Horejsi of September 30, 2021. and Tomas Brdicka J Immunol 2013; 190:1807-1818; Prepublished online 18 January 2013; doi: 10.4049/jimmunol.1202401 http://www.jimmunol.org/content/190/4/1807 Downloaded from Supplementary http://www.jimmunol.org/content/suppl/2013/01/18/jimmunol.120240 Material 1.DC1 http://www.jimmunol.org/ References This article cites 75 articles, 32 of which you can access for free at: http://www.jimmunol.org/content/190/4/1807.full#ref-list-1 Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision by guest on September 30, 2021 • No Triage! Every submission reviewed by practicing scientists • Fast Publication! 4 weeks from acceptance to publication *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2013 by The American Association of Immunologists, Inc. All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology Nonredundant Roles of Src-Family Kinases and Syk in the Initiation of B-Cell Antigen Receptor Signaling Ondrej Stepanek, Peter Draber, Ales Drobek, Vaclav Horejsi, and Tomas Brdicka When a BCR on a mature B cell is engaged by its ligand, the cell becomes activated, and the Ab-mediated immune response can be triggered. -

The Role of Signaling Pathways in the Development and Treatment of Hepatocellular Carcinoma
Oncogene (2010) 29, 4989–5005 & 2010 Macmillan Publishers Limited All rights reserved 0950-9232/10 www.nature.com/onc REVIEW The role of signaling pathways in the development and treatment of hepatocellular carcinoma S Whittaker1,2, R Marais3 and AX Zhu4 1Dana-Farber Cancer Institute, Boston, MA, USA; 2The Broad Institute, Cambridge, MA, USA; 3Institute of Cancer Research, London, UK and 4Massachusetts General Hospital Cancer Center, Harvard Medical School, Boston, MA, USA Hepatocellular carcinoma (HCC) is a highly prevalent, malignancy in adults (Pons-Renedo and Llovet, 2003). treatment-resistant malignancy with a multifaceted mole- For the vast majority of patients, HCC is a late cular pathogenesis. Current evidence indicates that during complication of chronic liver disease, and as such, is hepatocarcinogenesis, two main pathogenic mechanisms often associated with cirrhosis. The main risk factors for prevail: (1) cirrhosis associated with hepatic regeneration the development of HCC include infection with hepatitis after tissue damage caused by hepatitis infection, toxins B virus (HBV) or hepatitis C virus (HCV). Hepatitis (for example, alcohol or aflatoxin) or metabolic influ- infection is believed to be the main etiologic factor in ences, and (2) mutations occurring in single or multiple 480% of cases (Anzola, 2004). Other risk factors oncogenes or tumor suppressor genes. Both mechanisms include excessive alcohol consumption, nonalcoholic have been linked with alterations in several important steatohepatitis, autoimmune hepatitis, primary biliary cellular signaling pathways. These pathways are of cirrhosis, exposure to environmental carcinogens (parti- interest from a therapeutic perspective, because targeting cularly aflatoxin B) and the presence of various genetic them may help to reverse, delay or prevent tumorigenesis. -

Epigenetic Gene Regulation by Janus Kinase 1 in Diffuse Large B-Cell Lymphoma
Epigenetic gene regulation by Janus kinase 1 in diffuse large B-cell lymphoma Lixin Ruia,b,c,1,2, Amanda C. Drennanb,c,1, Michele Ceribellia,1, Fen Zhub,c, George W. Wrightd, Da Wei Huanga, Wenming Xiaoe, Yangguang Lib,c, Kreg M. Grindleb,c,LiLub,c, Daniel J. Hodsona, Arthur L. Shaffera, Hong Zhaoa, Weihong Xua, Yandan Yanga, and Louis M. Staudta,2 aLymphoid Malignancies Branch, Center for Cancer Research, National Cancer Institute, NIH, Bethesda, MD 20892; bDepartment of Medicine, School of Medicine and Public Health, University of Wisconsin, Madison, WI 53705; cCarbone Cancer Center, School of Medicine and Public Health, University of Wisconsin, Madison, WI 53705; dBiometric Research Branch, DCTD, National Cancer Institute, NIH, Bethesda, MD 20892; and eDivision of Bioinformatics and Biostatistics, National Center for Toxicological Research/Food and Drug Administration, Jefferson, AR 72079 Contributed by Louis M. Staudt, September 29, 2016 (sent for review July 22, 2016; reviewed by Anthony R. Green and Ross L. Levine) Janus kinases (JAKs) classically signal by activating STAT transcription promote STAT dimerization, nuclear translocation, and binding factors but can also regulate gene expression by epigenetically to cis-regulatory elements to regulate transcription (15, 17). This phosphorylating histone H3 on tyrosine 41 (H3Y41-P). In diffuse large canonical JAK/STAT pathway is deregulated in several hemato- B-cell lymphomas (DLBCLs), JAK signaling is a feature of the activated logic malignancies (16). In DLBCL, STAT3 is activated in the B-cell (ABC) subtype and is triggered by autocrine production of IL-6 ABC subtype and regulates gene expression to promote the sur- and IL-10. -

Activation Tyrosine Kinases, Btk and Lyn, in Mast Cell Redundant And
Redundant and Opposing Functions of Two Tyrosine Kinases, Btk and Lyn, in Mast Cell Activation This information is current as Yuko Kawakami, Jiro Kitaura, Anne B. Satterthwaite, of September 25, 2021. Roberta M. Kato, Koichi Asai, Stephen E. Hartman, Mari Maeda-Yamamoto, Clifford A. Lowell, David J. Rawlings, Owen N. Witte and Toshiaki Kawakami J Immunol 2000; 165:1210-1219; ; doi: 10.4049/jimmunol.165.3.1210 Downloaded from http://www.jimmunol.org/content/165/3/1210 References This article cites 75 articles, 45 of which you can access for free at: http://www.jimmunol.org/content/165/3/1210.full#ref-list-1 http://www.jimmunol.org/ Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists by guest on September 25, 2021 • Fast Publication! 4 weeks from acceptance to publication *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2000 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. Redundant and Opposing Functions of Two Tyrosine Kinases, Btk and Lyn, in Mast Cell Activation1 Yuko Kawakami,* Jiro Kitaura,* Anne B. Satterthwaite,† Roberta M. -

JAK3 Deficiency, (SCID T-B+)
JAK3 deficiency, (SCID T-B+) Author: Professor Luigi D. Notarangelo1,2 Creation Date: November 2001 Update: January 2005 1member of the European editorial committee of Orphanet encyclopedia 2Department of Pediatrics, University of Brescia, Spedali Civil, 25123 Brescia, Italy. [email protected] Abstract Keywords Diagnosis criteria/definition Differential diagnosis Prevalence Clinical description Treatment Etiology Diagnostic methods Genetic counseling Antenatal diagnosis Unresolved questions References Abstract JAK3 (Janus Kinase 3) deficiency is an autosomal recessive form of severe combined immune deficiency (SCID). It is characterized by lack of circulating T and NK (Natural Killer) cells and normal number of B lymphocytes. The disease is due to mutations in the JAK3 gene encoding an intracellular tyrosine kinase that is physically and functionally coupled with several cytokine receptors. Identification of gene anomalies has allowed physicians to make the diagnosis (even prenatal), and may prompt novel forms of treatment based on gene therapy. Although a relatively low number of JAK3-deficient subjects have been diagnosed, JAK3 deficiency represents an important cause of autosomal recessive SCID in the United States and its prevalence in Europe appears to be even higher. However it is considered as a rare disease (incidence is between 1/100,000 and 1/1,000,000 live births). JAK3-deficient patients present with the classical clinical features of SCID in the first few months of life, i.e. chronic diarrhea, failure to thrive, recurrent respiratory infection and/or generalized infections from opportunistic pathogens, or signs of graft-versus-host reaction (skin rash, abnormalities of liver function, pancytopenia) from transplacental acquired maternal T cells. -

Advances in Systemic Therapy for HER2-Positive Metastatic Breast Cancer Phuong Khanh H Morrow1, Francisco Zambrana1,2 and Francisco J Esteva1
Available online http://breast-cancer-research.com/content/11/4/207 Review Advances in systemic therapy for HER2-positive metastatic breast cancer Phuong Khanh H Morrow1, Francisco Zambrana1,2 and Francisco J Esteva1 1Department of Breast Medical Oncology, The University of Texas MD Anderson Cancer Center, Holcombe Boulevard, Houston, TX 77030, USA 2Current address: Department of Medical Oncology, Hospital Infanta Sofia, Paseo de Europa, 34, Madrid 28702, Spain Corresponding author: Phuong Khanh H Morrow, [email protected] Published: 15 July 2009 Breast Cancer Research 2009, 11:207 (doi:10.1186/bcr2324) This article is online at http://breast-cancer-research.com/content/11/4/207 © 2009 BioMed Central Ltd Abstract differentiation, survival, and migration that are associated with Human epidermal growth factor receptor (HER)2 over-expression HER2-positive breast cancers (Figure 1). Thus, women with is associated with a shortened disease-free interval and poor HER2-positive breast cancers exhibit significantly decreased survival. Although the addition of trastuzumab to chemotherapy in disease-free survival and overall survival (OS) [2-5]. the first-line setting has improved response rates, progression-free survival, and overall survival, response rates declined when trastu- This review discusses progress in the treatment of HER2- zumab was used beyond the first-line setting because of multiple positive metastatic breast cancer since the discovery of the mechanisms of resistance. Studies have demonstrated the clinical utility of continuing