What Is Stress? Concepts, Definitions and Applications in Seed Science
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Plant Physiology General the Main Light Sensitive Pigment Able to Absorb Solar Energy in Both Plants and Algae Is
Plant Physiology General the main light sensitive pigment able to absorb solar energy in both plants and algae is chlorophyll Photosynthesis this chlorophyll is contained with the chloroplasts probably the most characteristic “thing” that plants plants also have other “accessory pigments”: “do” is photosynthesis carotenoids – mainly yellow, orange almost all plants are autotrophs but usually their colors are masked by an abundance of !use energy from the sun to make sugar and chlorophyll other organic molecules out of simple fall colors are seen as a deciduous plant shuts down nutrients and chlorophyll is broken down and recycled leaving the colors of the other pigments photosynthesis requires carbon dioxide & water reds come from anthocyanins made to protect leaves as they recycle nutrients from the breakdown of chlorophyll CO2 enters through stomata or pores [Application] water is absorbed through roots researchers are studying the structure of the chloroplasts to light improve efficiency in the design of solar collectors CO2 + H2O sugar + O2 chlorophyll (glucose) today (2006) the most efficient solar cells capture only ~17% of solar energy that lands on them, while plant [photosynthesis converts water and carbon dioxide cell capture 30-40% to sugar and oxygen] !these sugars can then be broken down as needed for energy photosynthesis uses several chemical pigment to absorb the energy from sunlight Plants: Plant Physiology - General, Ziser, Lecture Notes, 2012.10 1 Plants: Plant Physiology - General, Ziser, Lecture Notes, 2012.10 2 Plant -

BIL 161: Environment and Development: the Effects of Environmental Variables on Seed Germination
BIL 161: Environment and Development: The Effects of Environmental Variables on Seed Germination The seed is more than just a plant waiting to happen. It is a complex marvel of evolution, a miniature life-support system that responds to environmental cues in order to give the embryo nestled within the best chance of survival. I. Characteristics and Classification of Plants Plants share synapomorphies that set them apart from other organisms. 1. true tissues (of types unique to plants) 2. waxy cuticle (to prevent desiccation) 3. stomates (microscopic gas exchange pores on the leaves) 4. apical meristems (permanent embryonic tissue for constant growth) 5. multicellular sex organs (male antheridia and female archegonia) 6. walled spores produced in structures called sporangia 7. embryo development inside the female parent 8. secondary metabolites (alkaloids, tannins, flavonoids, etc.) 9. heteromorphic alternation of generations The most primitive plants do not produce seeds at all, but rather release spores into the environment where they grow into a second life cycle stage, called the gametophyte. In seed plants, the life cycle is highly derived. Seed plants still make spores, but each spore grows into a gametophyte that is little more than a bit of tissue that gives rise to gametes. In the male parts of the plant, each spore develops into a sperm-producing male gametophyte known as pollen. In the female parts of the plant, meiosis occurs inside a structure known as the ovule, which will eventually give rise to the seed. Plants can broadly be classified as follows. A. Bryophytes – non-vascular plants (mosses, liverworts and hornworts) B. -

Plant Physiology
PLANT PHYSIOLOGY Vince Ördög Created by XMLmind XSL-FO Converter. PLANT PHYSIOLOGY Vince Ördög Publication date 2011 Created by XMLmind XSL-FO Converter. Table of Contents Cover .................................................................................................................................................. v 1. Preface ............................................................................................................................................ 1 2. Water and nutrients in plant ............................................................................................................ 2 1. Water balance of plant .......................................................................................................... 2 1.1. Water potential ......................................................................................................... 3 1.2. Absorption by roots .................................................................................................. 6 1.3. Transport through the xylem .................................................................................... 8 1.4. Transpiration ............................................................................................................. 9 1.5. Plant water status .................................................................................................... 11 1.6. Influence of extreme water supply .......................................................................... 12 2. Nutrient supply of plant ..................................................................................................... -

What Are Soybeans?
candy, cakes, cheeses, peanut butter, animal feeds, candles, paint, body lotions, biodiesel, furniture soybeans USES: What are soybeans? Soybeans are small round seeds, each with a tiny hilum (small brown spot). They are made up of three basic parts. Each soybean has a seed coat (outside cover that protects the seed), VOCABULARY cotyledon (the first leaf or pair of leaves within the embryo that stores food), and the embryo (part of a seed that develops into Cultivar: a variety of plant that has been created or a new plant, including the stem, leaves and roots). Soybeans, selected intentionally and maintained through cultivation. like most legumes, perform nitrogen fixation. Modern soybean Embryo: part of a seed that develops into a new plant, cultivars generally reach a height of around 1 m (3.3 ft), and including the stem, leaves and roots. take 80–120 days from sowing to harvesting. Exports: products or items that the U.S. sells and sends to other countries. Exports include raw products like whole soybeans or processed products like soybean oil or Leaflets soybean meal. Fertilizer: any substance used to fertilize the soil, especially a commercial or chemical manure. Hilum: the scar on a seed marking the point of attachment to its seed vessel (the brown spot). Leaflets: sub-part of leaf blade. All but the first node of soybean plants produce leaves with three leaflets. Legume: plants that perform nitrogen fixation and whose fruit is a seed pod. Beans, peas, clover and alfalfa are all legumes. Nitrogen Fixation: the conversion of atmospheric nitrogen Leaf into a nitrogen compound by certain bacteria, such as Stem rhizobium in the root nodules of legumes. -

International Union for the Protection of New Varieties of Plants
E TG/81/7(proj.1) ORIGINAL: English DATE: 2018-04-05 INTERNATIONAL UNION FOR THE PROTECTION OF NEW VARIETIES OF PLANTS Geneva DRAFT * COMMON SUNFLOWER UPOV Code(s): HLNTS_ANN Helianthus annuus L. GUIDELINES FOR THE CONDUCT OF TESTS FOR DISTINCTNESS, UNIFORMITY AND STABILITY prepared by experts from Hungary to be considered by the Technical Working Party for Agricultural Crops at its forty-seventh session, to be held in Naivasha, Kenya, from 2018-05-21 to 2018-05-25 Disclaimer: this document does not represent UPOV policies or guidance Alternative names:* Botanical name English French German Spanish Helianthus annuus L. Common Sunflower Soleil, Tournesol Sonnenblume Girasol The purpose of these guidelines (“Test Guidelines”) is to elaborate the principles contained in the General Introduction (document TG/1/3), and its associated TGP documents, into detailed practical guidance for the harmonized examination of distinctness, uniformity and stability (DUS) and, in particular, to identify appropriate characteristics for the examination of DUS and production of harmonized variety descriptions. ASSOCIATED DOCUMENTS These Test Guidelines should be read in conjunction with the General Introduction and its associated TGP documents. * These names were correct at the time of the introduction of these Test Guidelines but may be revised or updated. [Readers are advised to consult the UPOV Code, which can be found on the UPOV Website (www.upov.int), for the latest information.] TG/81/7(proj.1) Common Sunflower, 2018-04-05 2 TABLE OF CONTENTS PAGE 1. -

Effects of Root Pruning on Germinated Pecan Seedlings
PROPAGATION AND TISSUE CULTURE HORTSCIENCE 50(10):1549–1552. 2015. if pruning the roots shortly after germination stimulates lateral root formation. Similarly, little work has been attempted to consider Effects of Root Pruning on Germinated the effects of taproot pruning in young pecan seedlings back to different lengths on root Pecan Seedlings regeneration. The objective of this study 1 was to determine if root pruning at different Rui Zhang, Fang-Ren Peng , Pan Yan, Fan Cao, periods after seed germination affects pecan and Zhuang-Zhuang Liu seedling root and shoot growth and to evaluate College of Forestry, Nanjing Forestry University, 159 Longpan Road, different degrees of taproot pruning on root Nanjing 210037, China initiation. Dong-Liang Le Materials and Methods College of Forestry, Nanjing Forestry University, 159 Longpan Road, Nanjing 210037, China; and Nanjing Green Universe Pecan Science and The test was established at Lvzhou Pecan Research Station, Nanjing, China (lat. Technology Co. Ltd., 38 Muxuyuan Street, Nanjing 210007, China 32.05°N, long. 118.77°E). Open-pollinated Peng-Peng Tan seeds of the Chinese selection ‘Shaoxing’ were used as seed stock to produce seedlings. College of Forestry, Nanjing Forestry University, 159 Longpan Road, ‘Shaoxing’ had a small nut of very good Nanjing 210037, China quality with 50% percentage fill. The seeds averaged 30.4 mm in length, 20.9 mm in Additional index words. Carya illinoinensis, taproot, lateral root, shoot growth width, and 5.4 g in weight. Seeds were Abstract. Root systems of pecan trees are usually dominated by a single taproot with few collected on 7 Nov. -

Botany for Gardeners Offers a Clear Explanation of How Plants Grow
BotGar_Cover (5-8-2004) 11/8/04 11:18 AM Page 1 $19.95/ £14.99 GARDENING & HORTICULTURE/Reference Botany for Gardeners offers a clear explanation of how plants grow. • What happens inside a seed after it is planted? Botany for Gardeners Botany • How are plants structured? • How do plants adapt to their environment? • How is water transported from soil to leaves? • Why are minerals, air, and light important for healthy plant growth? • How do plants reproduce? The answers to these and other questions about complex plant processes, written in everyday language, allow gardeners and horticulturists to understand plants “from the plant’s point of view.” A bestseller since its debut in 1990, Botany for Gardeners has now been expanded and updated, and includes an appendix on plant taxonomy and a comprehensive index. Twodozen new photos and illustrations Botany for Gardeners make this new edition even more attractive than its predecessor. REVISED EDITION Brian Capon received a ph.d. in botany Brian Capon from the University of Chicago and was for thirty years professor of botany at California State University, Los Angeles. He is the author of Plant Survival: Adapting to a Hostile Brian World, also published by Timber Press. Author photo by Dan Terwilliger. Capon For details on other Timber Press books or to receive our catalog, please visit our Web site, www.timberpress.com. In the United States and Canada you may also reach us at 1-800-327-5680, and in the United Kingdom at [email protected]. ISBN 0-88192-655-8 ISBN 0-88192-655-8 90000 TIMBER PRESS 0 08819 26558 0 9 780881 926552 UPC EAN 001-033_Botany 11/8/04 11:20 AM Page 1 Botany for Gardeners 001-033_Botany 11/8/04 11:21 AM Page 2 001-033_Botany 11/8/04 11:21 AM Page 3 Botany for Gardeners Revised Edition Written and Illustrated by BRIAN CAPON TIMBER PRESS Portland * Cambridge 001-033_Botany 11/8/04 11:21 AM Page 4 Cover photographs by the author. -
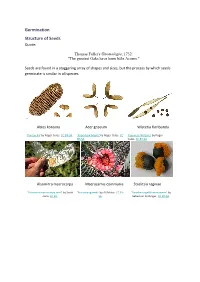
Germination Structure of Seeds Quote
Germination Structure of Seeds Quote: Thomas Fuller's Gnomologia, 1732: "The greatest Oaks have been little Acorns." Seeds are found in a staggering array of shapes and sizes, but the process by which seeds germinate is similar in all species. Abies koreana Acer griseum Wisteria floribunda 'Korean Fir' by Roger Culos. CC BY-SA. 'Paperbark Maple' by Roger Culos. CC 'Japanese Wisteria' by Roger BY-SA. Culos. CC BY-SA. Alsomitra macrocarpa Macrozamia communis Strelitzia reginae 'Alsomitra macrocarpa seed' by Scott 'BurrawangSeeds' by AYArktos. CC BY- 'Paradiesvogelblumensamen' by Zona. CC BY. SA. Sebastian Stabinger. CC BY-SA. Taraxicum officinale Stephanotis floribunda Phleum pratense 'Achane of Taraxacum sect. 'Stephanotis seed' by L. Marie"/Lenore 'Timoteegras vruchten Phleum Ruderalia' by Didier Edman, Sunnyvale, CA. CC BY. pratense' by Rasbak. CC BY-SA. Descouens. CC BY-SA. Dicotyledon seeds testa epicotyl plumule hypocotyl cotyledon radicle 'Aesculus hippocastanum seed section' by Boronian. CC BY. plumule epicotyl hypocotyl testa hilum radicle cotyledon micropyle endosperm Monocotyledon seeds endosperm epicotyl testa hypocotyl cotyledon radicle Parts of a seed Testa The seed coat. A protective layer which is tough and hard and it protects the seed from attack by insects, fungi and bacteria. Cotyledon Dicotyledons have 2 cotyledons Monocotyledons have 1 cotyledon A cotyledon is an embryonic leaf. It is the first leaf to appear when a seedling grows. They often contain reserves of food which the developing seedling can use to grow. Epicotyl The section of stem between the cotyledon(s) and the plumule. In a seedling it is the section of stem between the cotyledons and the first true leaves. -

Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris Richardii
G C A T T A C G G C A T genes Article Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii Alejandro Aragón-Raygoza 1,2 , Alejandra Vasco 3, Ikram Blilou 4, Luis Herrera-Estrella 2,5 and Alfredo Cruz-Ramírez 1,* 1 Molecular and Developmental Complexity Group at Unidad de Genómica Avanzada, Laboratorio Nacional de Genómica para la Biodiversidad, Cinvestav Sede Irapuato, Km. 9.6 Libramiento Norte Carretera, Irapuato-León, Irapuato 36821, Guanajuato, Mexico; [email protected] 2 Metabolic Engineering Group, Unidad de Genómica Avanzada, Laboratorio Nacional de Genómica para la Biodiversidad, Cinvestav Sede Irapuato, Km. 9.6 Libramiento Norte Carretera, Irapuato-León, Irapuato 36821, Guanajuato, Mexico; [email protected] 3 Botanical Research Institute of Texas (BRIT), Fort Worth, TX 76107-3400, USA; [email protected] 4 Laboratory of Plant Cell and Developmental Biology, Division of Biological and Environmental Sciences and Engineering (BESE), King Abdullah University of Science and Technology (KAUST), Thuwal 23955-6900, Saudi Arabia; [email protected] 5 Institute of Genomics for Crop Abiotic Stress Tolerance, Department of Plant and Soil Science, Texas Tech University, Lubbock, TX 79409, USA * Correspondence: [email protected] Received: 27 October 2020; Accepted: 26 November 2020; Published: 4 December 2020 Abstract: Ferns are a representative clade in plant evolution although underestimated in the genomic era. Ceratopteris richardii is an emergent model for developmental processes in ferns, yet a complete scheme of the different growth stages is necessary. Here, we present a developmental analysis, at the tissue and cellular levels, of the first shoot-borne root of Ceratopteris. -

Morphological Description of Plants: New Perspectives in Development and Evolution
® International Journal of Plant Developmental Biology ©2010 Global Science Books Morphological Description of Plants: New Perspectives in Development and Evolution 1* 2 Emilio Cervantes • Juana G . de Diego 1 IRNASA-CSIC. Apartado 257. 37080. Salamanca. Spain 2 Dept Bioquímica y Biología Molecular. Campus Miguel de Unamuno. Universidad de Salamanca. Spain Corresponding author : * [email protected] ABSTRACT The morphological description of plants has been fundamental in the history of botany and provided the keys for taxonomy. Nevertheless, in biology, a discipline governed by the interest in function and based on reductionist approaches, the analysis of forms has been relegated to second place. Plants contain organs and structures that resemble geometrical forms. Plant ontogeny may be seen as a sequence of growth processes including periods of continuous growth with modification that stop at crucial points often represented by structures of remarkable similarity to geometrical figures. Instead of the tradition in developmental studies giving more importance to animal models, we propose that the modular type of plant development may serve to remark conceptual aspects in that may be useful in studies with animals and contribute to original views of evolution. _____________________________________________________________________________________________________________ Keywords: concepts, description, geometry, structure INTRODUCTION: PLANTS OFFER NEW reflects a more general situation in Biology, a discipline APPROACHES TO DEVELOPMENT that has matured from the experimental protocols and views of biochemistry, where the predominant role of experimen- The study of development is today a major biological disci- tation has contributed to a decline in basic aspects that need pline. Although it was traditionally more focused in animal to be more descriptive, such as those related with mor- systems, increased emphasis in plant development may phology. -

Roper, Hemmons
ORIGINS OF TRANSLOCATIONS IN ASPERGZLLUS NZDULANSl ETTA KAFER Department of Genetics, McGill University, Montreal, Cam& T was realized early in the course of the genetic analysis of the ascomycete ‘Aspergillus nidukns that several of the strains with X-ray induced mutants contain chromosomal aberrations (e.g. two cases mentioned by PONTECORVO, ROPER,HEMMONS, MACDONALD and BUFTON1953). At that time all evidence came from meiotic analysis. In crosses heterozygous for chromosomal aberra- tions, crossing over appears to be reduced since certain unbalanced crossover types show low viability. In fungi, this results in an unusually high frequency of aborted and abnormal ascospores, corresponding to defective pollen in higher plants that show “semisterility” as a result of chromosome aberrations. Ascopore patterns can be used to detect visually the presence of aberrations in Neurospora, either in intact asci (MCCLINTOCK1945), or in random spores (PERKINS, GLASSEYand BLOOM1962). The nonlinear ascus of Aspergillus is too small to make use of this method routinely even though similar patterns have been ob- served in asci from a cross which is now known to have been heterozygous for at least three aberrations (ELLIOTT1960). On the other hand, the discovery of mitotic recombination in Aspergillus (ROPER1952; PONTECORVOand ROPER 1953) has provided new genetic methods not only for the mapping of markers but also for the detection of aberrations, especially translocations (PONTECORVO and =FER 1958; KAFER 1958). Even though most X-ray mutants had been excluded from the general Glasgow stocks, several aberrations have been encountered in the course of mitotic mapping of new markers. The first of these resulted in mitotic linkage between markers of linkage groups I and VII. -

Tree Seedling Availability, Planting, and Initial Care
F-5024 Tree Seedling Availability, Planting, and Initial Care William G. Ross Assistant Professor Oklahoma Cooperative Extension Fact Sheets Extension Forestry Specialistst are also available on our website at: http://www.osuextra.com Introduction This is the second in a series of three publications addressing the topic of forest stand establishment. If one is Bareroot not sure of what seedlings to plant or of what forest product to Most nurseries that sell seedlings produce some form of produce, it may be beneficial to read the first publication in this bareroot stock. Bareroot seedlings, as the name describes, is series, titled “Tree Planting Objectives and the Seedling a seedling with only the stem and the root supplied to the Selection Process.” landowner for planting. The major advantages of bareroot There are four points toconsider to identify management stock are the relative ease of production in the nurseries, the objectives. These are listed in brief form below. relative low cost to landowner, the relative ease of planting, and the ability to mechanize many of the operations in the 1. Identify the growing region where land is located. seedling production method. 2. Identify the soil types and moisture regimes located on the Shortcomings of bareroot seedlings include the vulner- land. ability of the scedling to the uncontrollable nursery environ- 3. Identify those objectives feasible and within the bounds of ment, exposure of the root system to harsh environmental the above environmental constraints. conditions after lifting, and the requirement of a large, sea- 4. Identify those tree species that, when managed correctly, sonal labor force to plant the seedlings.