Seminar: Modern Classics in Visual Perception, Visual Science, and Visual Neuroscience: Psychology W3255 Psychology G4255y
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Visual Cortex in Humans 251
Author's personal copy Visual Cortex in Humans 251 Visual Cortex in Humans B A Wandell, S O Dumoulin, and A A Brewer, using fMRI, and we discuss the main features of the Stanford University, Stanford, CA, USA V1 map. We then summarize the positions and proper- ties of ten additional visual field maps. This represents ã 2009 Elsevier Ltd. All rights reserved. our current understanding of human visual field maps, although this remains an active field of investigation, with more maps likely to be discovered. Finally, we Human visua l cortex comprises 4–6 billion neurons that are organ ized into more than a dozen distinct describe theories about the functional purpose and functional areas. These areas include the gray matter organizing principles of these maps. in the occi pital lobe and extend into the temporal and parietal lobes . The locations of these areas in the The Size and Location of Human Visual intact human cortex can be identified by measuring Cortex visual field maps. The neurons within these areas have a variety of different stimulus response proper- The entirety of human cortex occupies a surface area 2 ties. We descr ibe how to measure these visual field on the order of 1000 cm and ranges between 2 and maps, their locations, and their overall organization. 4 mm in thickness. Each cubic millimeter of cortex contains approximately 50 000 neurons so that neo- We then consider how information about patterns, objects, color s, and motion is analyzed and repre- cortex in the two hemispheres contain on the order of sented in these maps. -

Auditory Experience Controls the Maturation of Song Discrimination and Sexual Response in Drosophila Xiaodong Li, Hiroshi Ishimoto, Azusa Kamikouchi*
RESEARCH ARTICLE Auditory experience controls the maturation of song discrimination and sexual response in Drosophila Xiaodong Li, Hiroshi Ishimoto, Azusa Kamikouchi* Graduate School of Science, Nagoya University, Nagoya, Japan Abstract In birds and higher mammals, auditory experience during development is critical to discriminate sound patterns in adulthood. However, the neural and molecular nature of this acquired ability remains elusive. In fruit flies, acoustic perception has been thought to be innate. Here we report, surprisingly, that auditory experience of a species-specific courtship song in developing Drosophila shapes adult song perception and resultant sexual behavior. Preferences in the song-response behaviors of both males and females were tuned by social acoustic exposure during development. We examined the molecular and cellular determinants of this social acoustic learning and found that GABA signaling acting on the GABAA receptor Rdl in the pC1 neurons, the integration node for courtship stimuli, regulated auditory tuning and sexual behavior. These findings demonstrate that maturation of auditory perception in flies is unexpectedly plastic and is acquired socially, providing a model to investigate how song learning regulates mating preference in insects. DOI: https://doi.org/10.7554/eLife.34348.001 Introduction Vocal learning in infants or juvenile birds relies heavily on the early experience of the adult conspe- cific sounds. In humans, early language input is necessary to form the ability of phonetic distinction *For correspondence: and pattern detection in the phase of auditory learning (Doupe and Kuhl, 1999; Kuhl, 2004). [email protected] Because of the strong parallels between speech acquisition of humans and song learning of song- Competing interests: The birds, and the difficulties to investigate the neural mechanisms of human early auditory memory at authors declare that no cellular resolution, songbirds have been used as a predominant model in studying memory formation competing interests exist. -

The Davida Teller Award Lecture, 2016 Visual Brain Development: A
Journal of Vision (2017) 17(3):26, 1–24 1 The Davida Teller Award Lecture, 2016 Visual Brain Development: A review of ‘‘Dorsal Stream Vulnerability’’—motion, mathematics, amblyopia, actions, and attention # Janette Atkinson University College London, London, UK $ Research in the Visual Development Unit on ‘‘dorsal stream vulnerability’ (DSV) arose from research in two Introduction somewhat different areas. In the first, using cortical milestones for local and global processing from our neurobiological model, we identified cerebral visual Emerging cortical function and infant cortical impairment in infants in the first year of life. In the impairment second, using photo/videorefraction in population refractive screening programs, we showed that infant The work discussed at the beginning of this review spectacle wear could reduce the incidence of strabismus arose out of the first twenty years of my research with and amblyopia, but many preschool children, who had Oliver Braddick and our team in the Visual Develop- been significantly hyperopic earlier, showed visuo-motor ment Unit in Cambridge, particularly John Wattam- and attentional deficits. This led us to compare Bell and Shirley Anker (Atkinson, 2000). We began by developing dorsal and ventral streams, using sensitivity devising new methods, both behavioral (automated to global motion and form as signatures, finding deficits forced-choice preferential looking) and electrophysio- in motion sensitivity relative to form in children with logical (steady-state VEP/VERP—Visual Evoked Po- Williams syndrome, or perinatal brain injury in tential/Visual Event Related Potential) to measure the hemiplegia or preterm birth. Later research showed that normal visual capacities of infants such as acuity and this DSV was common across many disorders, both ‘‘ ’’ contrast sensitivity, over the first years of life (Atkin- genetic and acquired, from autism to amblyopia. -
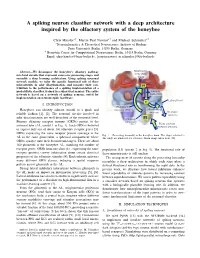
A Spiking Neuron Classifier Network with a Deep Architecture Inspired By
A spiking neuron classifier network with a deep architecture inspired by the olfactory system of the honeybee Chris Hausler¨ ∗†, Martin Paul Nawrot∗† and Michael Schmuker∗† ∗Neuroinformatics & Theoretical Neuroscience, Institute of Biology Freie Universitat¨ Berlin, 14195 Berlin, Germany † Bernstein Center for Computational Neuroscience Berlin, 10115 Berlin, Germany Email: [email protected], fmartin.nawrot, [email protected] Abstract—We decompose the honeybee’s olfactory pathway Mushroom into local circuits that represent successive processing stages and body (MB) calyx resemble a deep learning architecture. Using spiking neuronal network models, we infer the specific functional role of these microcircuits in odor discrimination, and measure their con- 2 tribution to the performance of a spiking implementation of a probabilistic classifier, trained in a supervised manner. The entire Kenyon network is based on a network of spiking neurons, suited for Cells (KC) implementation on neuromorphic hardware. Lateral horn 3 I. INTRODUCTION Projection Honeybees can identify odorant stimuli in a quick and neurons (PN) MB-extrinsic neurons To motor reliable fashion [1], [2]. The neuronal circuits involved in neurons odor discrimination are well described at the structural level. 1 Antennal Primary olfactory receptor neurons (ORNs) project to the lobe (AL) From odorant antennal lobe (AL, circuit 1 in Fig. 1). Each ORN is believed receptor neurons to express only one of about 160 olfactory receptor genes [3]. ORNs expressing the same receptor protein converge in the Fig. 1. Processing hierarchy in the honeybee brain. The stages relevant to AL in the same glomerulus, a spherical compartment where this study are numbered for reference. -

VISUAL NEUROSCIENCE Spring 2019
PSYC/BIBB/VLST 217 VISUAL NEUROSCIENCE Spring 2019 Spring 2019 Lecture: MWF 9-10a, Leidy Labs 10 Prerequisites: PSCY 1, BIBB 109, VLST 101, or COGS 001 Synopsis: An introduction to the scientific study of vision, with an emphasis on the biological substrate and its relation to behavior. Topics will typically include physiological optics, transduction of light, visual thresholds, color vision, anatomy and physiology of the visual pathways, and the cognitive neuroscience of vision. Instructor: Alan A. Stocker, Ph.D. Office: Goddard Labs Rm 421, 3710 Hamilton Walk Phone: (215) 573 9341 Email: [email protected] Office hour: W 12-1p Teaching Assistant: Lingqi Zhang Email: [email protected] Office hour: T 5-6p, TR 5-6p Office hour location: Goddard Labs Rm 420, 3710 Hamilton Walk Course Website (CANVAS): The course has a dedicated CANVAS site. Lecture slides, homework assignments, and reading assignments/material will be posted there. Also, check the site frequently for posted announcements and Q & As on the discussion board. In general, this is the place to go first if you have any question or are in need of any information regarding the course. Requirements: Homework1-3 posted on canvas Midterm Exam 1 February 15 (in class) Midterm Exam 2 March 29 (in class) Midterm Exam 3 April 26 (in class) Final Exam (cumulative) May 13, 12-2p Policy on homework assignments: Homework assignments are essential for a proper understanding of the material. There will be three homework assignments, meant in part as preparation for each midterm exam. Assignments and due dates will be posted on CANVAS. -

Perception, As You Make It
Perception, As You Make It Behavioral & Brain Sciences commentary on Chaz Firestone & Brian Scholl, “Cognition does not affect perception: Evaluating the evidence for ‘top-down’ effects" David W. Vinson, University of California, Merced, Drew H. Abney, University of California, Merced Dima Amso, Brown University Anthony Chemero, University of Cincinnati James E. Cutting, Cornell University Rick Dale, University of California, Merced Jonathan B. Freeman, New York University Laurie B. Feldman, The University at Albany, SUNY Karl J. Friston, University College London Shaun Gallagher, University of Memphis J. Scott Jordan, Illinois State University Liad Mudrik, Tel Aviv University Sasha Ondobaka, University College London Daniel C. Richardson, University College London Ladan Shams, University of California, Los Angeles Maggie Shiffrar, California State University, Northridge Michael J. Spivey, University of California, Merced Abstract: The main question F&S pose is whether “what and how we see is functionally independent from what and how we think, know, desire, act, etc.” We synthesize a collection of concerns from an interdisciplinary set of co-authors regarding F&S’s assumptions and appeals to intuition, resulting in their treatment of visual perception as context-free. No perceptual task takes place in a contextual vacuum. How do we know that an effect is one of perception qua perception that does not involve other cognitive contributions? Experimental instructions alone involve various cognitive factors that guide task performance (Roepstorff & Frith, 2004). Even a request to detect simple stimulus features requires participants to understand the instructions (“language, memory”), keep track of them (“working memory”), become sensitive to them (“attention”), and pick up the necessary information to become appropriately sensitive (“perception”). -

1P MON. PM Cortex and Subcortical Auditory Nuclei
dynamic role for the vestibular system in orientation and flight control. Laboratory studies of flight behavior under illuminated and dark conditions in both static and rotating obstacle tests were carried out while administering heavy water ͑D2O͒ to bats to impair their vestibular inputs. Eptesicus carried out complex maneuvers through both fixed arrays of wires and a rotating obstacle array using both vision and echolocation, or when guided by echolocation alone. When treated with D2O in combination with lack of visual cues, bats showed considerable decrements in performance. These data indicate that big brown bats use both vision and echolocation to provide spatial registration for head position information generated by the vestibular system. 2:55 1pAB3. Bat’s auditory system: Corticofugal feedback and plasticity. Nobuo Suga ͑Dept. of Biol., Washington Univ., One Brookings Dr., St. Louis, MO 63130͒ The auditory system of the mustached bat consists of physiologically distinct subdivisions for processing different types of biosonar information. It was found that the corticofugal ͑descending͒ auditory system plays an important role in improving and adjusting auditory signal processing. Repetitive acoustic stimulation, cortical electrical stimulation or auditory fear conditioning evokes plastic changes of the central auditory system. The changes are based upon egocentric selection evoked by focused positive feedback associated with lateral inhibition. Focal electric stimulation of the auditory cortex evokes short-term changes in the auditory 1p MON. PM cortex and subcortical auditory nuclei. An increase in a cortical acetylcholine level during the electric stimulation changes the cortical changes from short-term to long-term. There are two types of plastic changes ͑reorganizations͒: centripetal best frequency shifts for expanded reorganization of a neural frequency map and centrifugal best frequency shifts for compressed reorganization of the map. -

Principles of Image Representation in Visual Cortex
In: The Visual Neurosciences, L.M. Chalupa, J.S. Werner, Eds. MIT Press, 2003: 1603-1615. Principles of Image Representation in Visual Cortex Bruno A. Olshausen Center for Neuroscience, UC Davis [email protected] 1 Introduction data. Neuroscientists are interested in understand- ing how the cortex extracts certain properties of the The visual cortex is responsible for most of our con- visual environment—surfaces, objects, textures, mo- scious perception of the visual world, yet we remain tion, etc.—from the data stream coming from the largely ignorant of the principles underlying its func- retina. Similarly, engineers are interested in design- tion despite progress on many fronts of neuroscience. ing algorithms capable of extracting structure con- The principal reason for this is not a lack of data, tained in images or sound—for example, to identify but rather the absence of a solid theoretical frame- and label parts within the body from medical imag- work for motivating experiments and interpreting ing data. These problems at their core are one in the findings. The situation may be likened to trying same, and progress in one domain will likely lead to to understand how birds fly without knowledge of new insights in the other. the principles of aerodynamics: no amount of ex- The key difficulty faced by both sides is that the perimentation or measurements made on the bird core principles of pattern analysis are not well un- itself will reveal the secret. The key experiment— derstood. No amount of experimentation or techno- measuring air pressure above and below the wing as logical tinkering alone can overcome this obstacle. -

Krogh's Principle
Introduction to Neuroscience: Behavioral Neuroscience Neuroethology, Comparative Neuroscience, Natural Neuroscience Nachum Ulanovsky Department of Neurobiology, Weizmann Institute of Science 2017-2018, 2nd semester Principles of Neuroethology Neuroethology seeks to understand the mechanisms by which the Neurobiology central nervous system controls the Neuroethology Ethology natural behavior of animals. • Focus on Natural behaviors: Choosing to study a well-defined and reproducible yet natural behavior (either Innate or Learned behavior) • Need to study thoroughly the animal’s behavior, including in the field: Neuroethology starts with a good understanding of Ethology. • If you study the animals in the lab, you need to keep them in conditions as natural as possible, to avoid the occurrence of unnatural behaviors. • Krogh’s principle 1 Krogh’s principle August Krogh Nobel prize 1920 “For such a large number of problems there will be some animal of choice or a few such animals on which it can be most conveniently studied. Many years ago when my teacher, Christian Bohr, was interested in the respiratory mechanism of the lung and devised the method of studying the exchange through each lung separately, he found that a certain kind of tortoise possessed a trachea dividing into the main bronchi high up in the neck, and we used to say as a laboratory joke that this animal had been created expressly for the purposes of respiration physiology. I have no doubt that there is quite a number of animals which are similarly "created" for special physiological -

Lateral Inhibition Effects Demonstrate That the Maturational State of Neurons That Then Inhibit Their Neighbors
522 Lateral Inhibition effects demonstrate that the maturational state of neurons that then inhibit their neighbors. When a the learner’s brain is crucial for the attainment of stimulus (such as a bar of light or any other stim- a language system. ulus) excites a number of neurons in the network Several questions remain open about the mecha- (in this case neurons 4e, 5e, 6e, and 7e), the effect nisms underlying language learning. One issue is of inhibition is to suppress the neurons just out- whether specific aspects of language acquisition side the edge of the bar (3e and 8e) because those should be attributed to language-specific versus neurons are inhibited but not excited. Further, general-purpose learning mechanisms. Another issue because the neurons just inside the edges of the is whether children’s native language can affect the bar (4e and 7e) are excited by light and only inhib- way they think, and whether language is necessary ited by one neighbor, they are especially active. or helpful for the development of human concepts. This leads to perceptual contrast enhancement at borders. Further research showed that lateral inhi- Anna Papafragou bition also applied to overlapping stimuli, and that its strength fell off with distance between the See also Aphasias; Audition: Cognitive Influences; Context Effects in Perception; Speech Perception; Top- interacting stimuli. Down and Bottom-Up Processing; Word Recognition Haldan Keffer Hartline won the Nobel Prize in 1967 for discovering lateral inhibition and its neu- ral correlates. The first inhibitory circuit in the Further Readings nervous system was found in the horseshoe crab (Limulus polyphemus). -

Sensory Receptors A17 (1)
SENSORY RECEPTORS A17 (1) Sensory Receptors Last updated: April 20, 2019 Sensory receptors - transducers that convert various forms of energy in environment into action potentials in neurons. sensory receptors may be: a) neurons (distal tip of peripheral axon of sensory neuron) – e.g. in skin receptors. b) specialized cells (that release neurotransmitter and generate action potentials in neurons) – e.g. in complex sense organs (vision, hearing, equilibrium, taste). sensory receptor is often associated with nonneural cells that surround it, forming SENSE ORGAN. to stimulate receptor, stimulus must first pass through intervening tissues (stimulus accession). each receptor is adapted to respond to one particular form of energy at much lower threshold than other receptors respond to this form of energy. adequate (s. appropriate) stimulus - form of energy to which receptor is most sensitive; receptors also can respond to other energy forms, but at much higher thresholds (e.g. adequate stimulus for eye is light; eyeball rubbing will stimulate rods and cones to produce light sensation, but threshold is much higher than in skin pressure receptors). when information about stimulus reaches CNS, it produces: a) reflex response b) conscious sensation c) behavior alteration SENSORY MODALITIES Sensory Modality Receptor Sense Organ CONSCIOUS SENSATIONS Vision Rods & cones Eye Hearing Hair cells Ear (organ of Corti) Smell Olfactory neurons Olfactory mucous membrane Taste Taste receptor cells Taste bud Rotational acceleration Hair cells Ear (semicircular -

Do We Know What the Early Visual System Does?
The Journal of Neuroscience, November 16, 2005 • 25(46):10577–10597 • 10577 Mini-Symposium Do We Know What the Early Visual System Does? Matteo Carandini,1 Jonathan B. Demb,2 Valerio Mante,1 David J. Tolhurst,3 Yang Dan,4 Bruno A. Olshausen,6 Jack L. Gallant,5,6 and Nicole C. Rust7 1Smith-Kettlewell Eye Research Institute, San Francisco, California 94115, 2Departments of Ophthalmology and Visual Sciences, and Molecular, Cellular, and Developmental Biology, University of Michigan, Ann Arbor, Michigan 48105, 3Department of Physiology, University of Cambridge, Cambridge CB2 1TN, United Kingdom, Departments of 4Molecular and Cellular Biology and 5Psychology and 6Helen Wills Neuroscience Institute and School of Optometry, University of California, Berkeley, Berkeley, California 94720, and 7Center for Neural Science, New York University, New York, New York 10003 We can claim that we know what the visual system does once we can predict neural responses to arbitrary stimuli, including those seen in nature. In the early visual system, models based on one or more linear receptive fields hold promise to achieve this goal as long as the models include nonlinear mechanisms that control responsiveness, based on stimulus context and history, and take into account the nonlinearity of spike generation. These linear and nonlinear mechanisms might be the only essential determinants of the response, or alternatively, there may be additional fundamental determinants yet to be identified. Research is progressing with the goals of defining a single “standard model” for each stage of the visual pathway and testing the predictive power of these models on the responses to movies of natural scenes.