This Article Appeared in a Journal Published by Elsevier. the Attached
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lake Champlain Alewife Impacts February 14, 2006 Workshop Summary
Lake Champlain Alewife Impacts February 14, 2006 Workshop Summary Native smelt (top) and alewife from Lake Champlain LCSG-05-06 Planning Committee: J. Ellen Marsden, Univ. of Vermont, Burlington, VT Eric Palmer, VTFW, Waterbury, VT Bill Schoch, NYSDEC, Ray Brook, NY Dave Tilton, USFWS, Essex Junction, VT Lisa Windhausen, LCBP, Grand Isle, VT Mark Malchoff, LCSG/SUNY Plattsburgh LCRI, Plattsburgh, NY Lake Champlain Sea Grant 101 Hudson Hall, Plattsburgh State University of NY 101 Broad Street Plattsburgh, NY 12901-2681 http://research.plattsburgh.edu/ LakeChamplainSeaGrantAquatics/ans.htm and Lake Champlain Basin Program 54 West Shore Road - Grand Isle, VT 05458 http://www.lcbp.org/ 2 To Alewife Workshop Participants and Interested Parties: August 23, 2006 Alewives are native to the Atlantic coast and typically spawn in freshwater rivers and lakes. They are commonly used as bait and have become established in many lakes across the United States following intentional introductions and accidental bait-bucket releases. Once established in a new waterbody, alewives can cause tremendous changes to a lake ecosystem. Alewives first ap- peared in Lake Champlain’s Missisquoi Bay in 2003; they appeared in the Northeast Arm and the Main Lake segments in 2004 and 2005. Alewives are well established in Lake St. Catherine, which drains to Lake Champlain 80+ miles south of the 2004 discovery point. Based on experiences in other states, it is believed that an alewife infestation in Lake Champlain could have substantial eco- nomic and ecological impacts. Because the specific impacts of a widespread alewife infestation on Lake Champlain are uncertain, Lake Champlain Sea Grant and the Lake Champlain Basin Program organized a workshop on February 14, 2006 to learn from resource managers and scientists with experience in the Great Lakes and Finger Lakes of New York. -
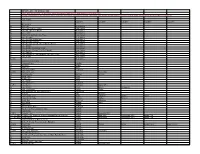
IMPORTANT INFORMATION: Lakes with an Asterisk * Do Not Have Depth Information and Appear with Improvised Contour Lines County Information Is for Reference Only
IMPORTANT INFORMATION: Lakes with an asterisk * do not have depth information and appear with improvised contour lines County information is for reference only. Your lake will not be split up by county. The whole lake will be shown unless specified next to name eg (Northern Section) (Near Follette) etc. LAKE NAME COUNTY COUNTY COUNTY COUNTY COUNTY Great Lakes GL Lake Erie Great Lakes GL Lake Erie (Port of Toledo) Great Lakes GL Lake Erie (Western Basin) Great Lakes GL Lake Huron Great Lakes GL Lake Huron (w West Lake Erie) Great Lakes GL Lake Michigan Great Lakes GL Lake Michigan (Northeast) Great Lakes GL Lake Michigan (South) Great Lakes GL Lake Michigan (w Lake Erie and Lake Huron) Great Lakes GL Lake Ontario Great Lakes GL Lake Ontario (Rochester Area) Great Lakes GL Lake Ontario (Stoney Pt to Wolf Island) Great Lakes GL Lake Superior Great Lakes GL Lake Superior (w Lake Michigan and Lake Huron) Great Lakes GL Lake St Clair Great Lakes GL (MI) Great Lakes Cedar Creek Reservoir AL Deerwood Lake Franklin AL Dog River Shelby AL Gantt Lake Mobile AL Goat Rock Lake * Covington AL (GA) Guntersville Lake Lee Harris (GA) AL Highland Lake * Marshall Jackson AL Inland Lake * Blount AL Jordan Lake Blount AL Lake Gantt * Elmore AL Lake Jackson * Covington AL (FL) Lake Martin Covington Walton (FL) AL Lake Mitchell Coosa Elmore Tallapoosa AL Lake Tuscaloosa Chilton Coosa AL Lake Wedowee (RL Harris Reservoir) Tuscaloosa AL Lay Lake Clay Randolph AL Lewis Smith Lake * Shelby Talladega Chilton Coosa AL Logan Martin Lake Cullman Walker Winston AL Mobile Bay Saint Clair Talladega AL Ono Island Baldwin Mobile AL Open Pond * Baldwin AL Orange Beach East Covington AL Bon Secour River and Oyster Bay Baldwin AL Perdido Bay Baldwin AL (FL) Pickwick Lake Baldwin Escambia (FL) AL (TN) (MS) Pickwick Lake (Northern Section, Pickwick Dam to Waterloo) Colbert Lauderdale Tishomingo (MS) Hardin (TN) AL (TN) (MS) Shelby Lakes Colbert Lauderdale Tishomingo (MS) Hardin (TN) AL Tallapoosa River at Fort Toulouse * Baldwin AL Walter F. -

A Revised Lake Trout Rehabilitation Plan for Ontario Waters of Lake Huron
A Revised Lake Trout Rehabilitation Plan for Ontario Waters of Lake Huron Upper Great Lakes Management Unit - Lake Huron 1 Upper Great Lakes Management Unit, Lake Huron Office MR-LHA- A Revised Lake Trout Rehabilitation Plan for Ontario Waters of Lake Huron May 4, 2012 Ontario Ministry of Natural Resources Upper Great Lakes Management Unit, Lake Huron Office 2 ©2012, Queen’s Printer for Ontario Printed in Ontario, Canada Upper Great Lakes Management Unit, Lake Huron Office Ministry of Natural Resources 1450 7th Avenue East Owen Sound, Ontario N4K 2Z1 (519) 371-0420 3 TABLE OF CONTENTS LIST OF FIGURES.............................................................................................. III LIST OF TABLES................................................................................................ III INTRODUCTION ................................................................................................... 1 WHY REHABILITATE LAKE TROUT? ............................................................................. 2 OVERVIEW OF PAST REHABILITATION EFFORTS .................................................... 3 SYNOPSIS OF PROGRESS TOWARDS LAKE TROUT REHABILITATION IN ONTARIO WATERS ............................................................................................................... 5 REVIEW OF THE 1996 PLAN ............................................................................................... 7 A REVISED GOAL AND OBJECTIVES FOR LAKE TROUT REHABILITION.................................................................................................... -

Life History and Reproductive Ecology of a Mid-Winter Spawner: the Burbot (Lota Lota)
Life history and reproductive ecology of a mid-winter spawner: the Burbot (Lota lota) by Peter Anton Cott Thesis submitted in partial fulfillment of the requirement for the degree of Doctor of Philosophy (PhD) in Boreal Ecology School of Graduate Studies Laurentian University Sudbury, Ontario © Peter Anton Cott, 2013 Library and Archives Bibliotheque et Canada Archives Canada Published Heritage Direction du 1+1 Branch Patrimoine de I'edition 395 Wellington Street 395, rue Wellington Ottawa ON K1A0N4 Ottawa ON K1A 0N4 Canada Canada Your file Votre reference ISBN: 978-0-494-92850-9 Our file Notre reference ISBN: 978-0-494-92850-9 NOTICE: AVIS: The author has granted a non L'auteur a accorde une licence non exclusive exclusive license allowing Library and permettant a la Bibliotheque et Archives Archives Canada to reproduce, Canada de reproduire, publier, archiver, publish, archive, preserve, conserve, sauvegarder, conserver, transmettre au public communicate to the public by par telecommunication ou par I'lnternet, preter, telecommunication or on the Internet, distribuer et vendre des theses partout dans le loan, distrbute and sell theses monde, a des fins commerciales ou autres, sur worldwide, for commercial or non support microforme, papier, electronique et/ou commercial purposes, in microform, autres formats. paper, electronic and/or any other formats. The author retains copyright L'auteur conserve la propriete du droit d'auteur ownership and moral rights in this et des droits moraux qui protege cette these. Ni thesis. Neither the thesis nor la these ni des extraits substantiels de celle-ci substantial extracts from it may be ne doivent etre imprimes ou autrement printed or otherwise reproduced reproduits sans son autorisation. -

Lake Trout Report
A. LAKE TROUT REHABILITATION GUIDE FOR LAKE HURON edited by Mark P. Ebener Inter-Tribal Fisheries and Assessment Program Chippewa/Ottawa Treaty Fishery Management Authority 179 W. 3 Mile Road Sault Ste. Marie, MI 49783 Citation: Ebener, M. P. [ED.]. 1998. A lake trout rehabilitation guide for Lake Huron. Great Lakes Fish. Comm. 48 p. Great Lakes Fishery Commission 2100 Commonwealth Blvd., Suite 209 Ann Arbor, MI 481051563 August 1998 TABLE OF CONTENTS ABSTRACT . ..1 INTRODUCTION ........................................................................................................................2 Background ........................................................................................................................3 MILESTONES AND INDICATORS . 5 INSIGHTS FROM RESTORE ....................................................................................................8 Stocking Strategies ............................................................................................................8 Population Regulation .......................................................................................................8 Community Regulation....................................................................................................10 Strategic Planning and Stockholder Involvement.. .......................................................... 10 REHABILITATION STRATEGIES ........................................................................................11 Rehabilitation Zones ........................................................................................................11 -

Indiana Aquatic Invasive Species (AIS) Management Plan 2020
Indiana Aquatic Invasive Species (AIS) Management Plan 2020 Indiana Department of Natural Resources, Division of Fish & Wildlife diana Dep,artment of Natural Resources i Table of Contents Executive Summary ..................................................................................................................... iii 1. Introduction ........................................................................................................................ 1 1.1 Aquatic Invasive Species Management Plan Purpose...................................................... 1 1.2 Aquatic Invasive Species Management Plan Background ............................................... 2 1.3 Past Accomplishments ..................................................................................................... 5 2. Problem and Prioritization ............................................................................................... 6 2.1 Animals .......................................................................................................................... 12 2.2 Plants .............................................................................................................................. 18 2.3 Diseases, Pathogens, and Parasites ................................................................................ 22 3. Existing Authorities and Programs ................................................................................ 25 3.1 Federal Role .................................................................................................................. -

219819Fre.Pdf
I I I I Le Programme federal des ports constitues en societe, les adminis Urgences pour petits bateaux trations portuaires sont dirigees par des Generatites : Plus de 2 200 ports pour petits bateaux personnes nommees par les groupes Centre de coordination des secours qui desservent une multitude de de pecheurs ou les collectivites. (Trenton) 1-800-267-7270 bateaux de peche commerciale, de En vertu d'une entente de cinq (Secours Air-Mer) - 24 heures sur 24 plaisance et autres, d'un ocean a l'autre, ans, ces administrations assument 1-613-267-7270 jouent un role vital dans la vie l'entretien courant de leurs ports. De pastes 3870 et 3875 economique et sociale du Canada. plus, elles etablissent et pergoivent les La responsabilite de l'entretien frais de location et les droits de permis Transport maritime - Situations permanent et de !'exploitation et d'amarrage. Ces recettes servent a d'urgence et Centre d'intervention en quotidienne de ces ports incombe au exploiter le port, a apporter certains cas de deversement (416) 268-6060 ministere federal des Peches et des niveaux precis de reparations et a Radio-urgence - Canal 16 156,80 kHz Oceans dans le cadre de son embaucher un responsable de port. Les Programme des ports pour petits projets d'entretien trop coOteux ou MAFORS (Meteo et changements pour bateaux (PPB). complexes pour !'administration la navigation) Canal 21 161 ,64 kHz Qu'il soit grand ou petit, un port est portuaire sont entrepris par les PPB . traditionnellement le centre d'activite Les ports geres par des adminis Les bateaux de sauvetage de la ainsi que l'assise economique de la trations portuaires ont la priorite pour ce Garde·cotiere plupart des collectivites de peche qui est du Programme des ports pour (du 24 mai au I er septembre) c6tieres. -

Bathymetry of Lake Huron with Topography
Bathymetry of Lake Huron with Topography 84ºW 80ºW 83ºW 82ºW 81ºW Lake Wakwekobi Lake Nipissing St. Joseph MICHIGAN Island Whitefish Falls 25 DrummondNorth Basin Channel 50 25 46ºN Hessel Drummond La Cloche 46ºN De Tour Drummond Peninsula Village Barrie Island French Basin Little River Current Cockburn Barrie ONTARIO Island Island Gore 25 Bay Kagawong Key Harbour 25 Thir Bayfield Soun t 50 d Kagawong Lake Lake Manitou y Wolsey moya Wikwemikong T 75 e Bois Blanc d L h 100 Lake in a Island k e o M u Byng Inlet s Duck Manitoulin a D Providence Bay Manitowaning n 125 Islands u d Cheboygan Co c Island ckbu k I rn s 25 Bas R South Bay l in South Baymouth i a d n g d e 50 Mullett Lake s Black Lake Huron 75 Beach Fitzwilliam 100 Island G e Brooks Landing Rogers City or gi an Parry Sound 125 Bruce B Presque Isle a Parry C y Island Manitoulin 150 Basin o l Tobermory l i 75 n Rockport 50 100 T 75 Basin175 g o B r 25 u w c b e 1 200 5 0 P o 125 e e n o 150 in Alpena r 100 s d m u 25 l 50 230 a C Lake Muskoka 100 50 l 200 150 45ºN o e h 45ºN Stokes Bay n r n Thunder Lions a y Head a n Bay A h l C n p Kn e o n e n ll e l s w Ossineke Th a - Pike Bay O un A d m Christian e Island Black r b River R id e Port Severn g r 100 Penetanguishene e l 75 Waubaushene e Thornbury Midland Victoria Harbour T H Port McNicoll h Wiarton u ar y n ris d vil R S Basin e le B i o r as d u B in t Harrisville a g h 50 si e a n m Meaford p 25 to n Owen Sound Thornbury O Harrisville B s a c s o i Collingwood d Knolls n Southampton a R 75 i d Oscoda g A Barrie e u s t i M n O i n Lake Simcoe c h t R a i g r i i Bay City a o d n g s Basin e East Tawa Tawas City dge Ri n G Alabaster l o o e r n u d Kincardine n a H e h 50 r C i c y t h i T OF E N C O M M T M C R E B A R - P C U.S. -

Fall 2014 Newsletter
“Wind Swept” November 2014 What’s inside 2014 LMAA information night World’s Greatest Lakes Culinary Delights Lightning Strikes Rocks of Lake Manitou Lessons from Geese Member’s Notice Board Director’s Roster LMAA information night 2014 by Dave Anderson Place: Sandfield School House Date: Tuesday, August 12, 2014 Time: 7:30 – 9:30 pm Agenda Introductions Invading species update by Eric Labelle Lyme Disease presentation by Helke Ferrie Meet your neighbour / refreshment break Blue Jay Creek Fish Culture update by Paul Methner Door prize Draws Approximately 55 hearty people ventured outside on a cool rainy night to attend the meeting at the Sandfield School house. Eric Labelle who was working on the Island for the Ontario Federation of Anglers and Hunters as part of the Invading Species Awareness Program gave an update on the invading species in and around Lake Manitou. The invading species program now has an early detection and distribution mapping system (EDDMaps) that allows quick and easy 1 “Wind Swept” November 2014 reporting of invasive species. Of course you may still report by telephone by calling 1-800-563-7711. Eric stressed the importance of being educated as to the types of invaders, and taking every precaution to prevent their entry into Lake Manitou and onto Manitoulin. Helke Ferrie, who is a medical journalist, gave an eye opening presentation on Lyme Disease in Ontario. Lyme Disease is a disease that is transmitted through the bite of a tick which are relatively common in natural areas. The disease is difficult to diagnose and treat and there is currently no vaccine to prevent its spread. -

Botanist Interior 43.1
2009 THE MICHIGAN BOTANIST 97 RARE PLANT SPECIES AND PLANT COMMUNITY TYPES OF MANITOU ISLAND AND GULL ROCK, KEWEENAW COUNTY, MICHIGAN Janet Keeney Marr Michael R. Penskar 23180 Highway Location Road Michigan Natural Features Inventory Calumet, MI 49913 Michigan State University Extension Stevens T. Mason Building, PO Box 30444 Lansing, MI 48909-7944 Dennis A. Albert 4073 Agriculture and Life Sciences Building Horticulture Department Oregon State University Corvallis, OR 97331-4501 ABSTRACT Botanical and ecological surveys of Manitou Island and Gull Rock were conducted in 1996 and 2005 to relocate and update the status of rare species associated with volcanic bedrock shoreline, as well as search for additional rare vascular plant species not previously known for Manitou Is - land. An additional objective of 2005 surveys was to search the interior of Manitou Island for rare plants, and to more comprehensively classify and describe the vegetation types of Manitou Island. Manitou Island is located in Lake Superior about 5 km east of the tip of the Keweenaw Penin - sula in Keweenaw County, Michigan. Manitou Island’s shoreline is comprised primarily of cobble beaches, steep sloping bedrock, and cliffs. The island’s bedrock shoreline has one of the highest concentrations of rare plant species for this natural community type in Michigan. One state en - dangered, nine state threatened, and four special concern species, several of which have boreal and arctic affinities, are now documented for Manitou Island, including two newly documented species, northern reedgrass ( Calamagrostis lacustris ) and fir club-moss ( Huperzia selago ), dis - covered in the island’s interior during the recent 2005 surveys. -

2014 Lake Erie Annual Report
NYS DEC LAKE ERIE 2014 ANNUAL REPORT to the Lake Erie Committee and the Great Lakes Fishery Commission March 2015 New York State Department of Environmental Conservation 625 Broadway, Albany, New York 12233-4753 Andrew M. Cuomo, Governor MOST OF THE WORK REPORTED IN THIS DOCUMENT IS SUPPORTED BY THE FEDERAL AID IN SPORT FISH RESTORATION PROGRAM Full Report Citation: NYSDEC 2015. Lake Erie 2014 Annual Report. New York State Department of Environmental Conservation, Albany, New York, USA Example Report Section Citation: Markham, J.L. 2015. Coldwater gill net assessment. Section F in NYSDEC 2015, Lake Erie 2014 Annual Report. New York State Department of Environmental Conservation, Albany, New York, USA NYS DEC LAKE ERIE 2014 ANNUAL REPORT to the Great Lakes Fishery Commission’s Lake Erie Committee NYSDEC Lake Erie Fisheries Research Unit e-mail: [email protected] Donald Einhouse, Lake Erie Unit Leader James Markham, Aquatic Biologist Jason Robinson, Aquatic Biologist Douglas Zeller, Research Vessel Captain Brian Beckwith, Fisheries Technician Richard Zimar, Fisheries Technician MariEllen (Ginger) Szwejbka, Secretary Steven LaPan, Great Lakes Fisheries Section Head Phil Hulbert, Chief, Bureau of Fisheries Acknowledgments The Lake Erie Fisheries Unit recognizes the contributions of seasonal staff essential to completing an ambitious field schedule. During the 2014 field year these individuals included Fish and Wildlife Technicians Carrie Babcock, Jonathon Draves, Ann Wilcox-Swanson, Kyle Keys and Robert Lichorat. We also acknowledge contributions of DEC’s Region 9 Fisheries Office, and Buffalo State College’s Great Lakes Center in support of various Lake Erie field activities. March 2015 New York State Department of Environmental Conservation 625 Broadway, Albany, New York 12233-4753 Preface The Lake Erie Annual Report is prepared by New York State Department of Environmental Conservation as a compilation of ongoing Lake Erie investigations mostly supported by Federal Aid in Sportfish Restoration. -

The State of Lake Huron in 1999 Special Publication 05-02
THE STATE OF LAKE HURON IN 1999 SPECIAL PUBLICATION 05-02 The Great Lakes Fishery Commission was established by the Convention on Great Lakes Fisheries between Canada and the United States, which was ratified on October 11, 1955. It was organized in April 1956 and assumed its duties as set forth in the Convention on July 1, 1956. The Commission has two major responsibilities: first, develop coordinated programs of research in the Great Lakes, and, on the basis of the findings, recommend measures which will permit the maximum sustained productivity of stocks of fish of common concern; second, formulate and implement a program to eradicate or minimize sea lamprey populations in the Great Lakes. The Commission is also required to publish or authorize the publication of scientific or other information obtained in the performance of its duties. In fulfillment of this requirement the Commission publishes the Technical Report Series, intended for peer-reviewed scientific literature; Special Publications, designed primarily for dissemination of reports produced by working committees of the Commission; and other (non-serial) publications. Technical Reports are most suitable for either interdisciplinary review and synthesis papers of general interest to Great Lakes fisheries researchers, managers, and administrators, or more narrowly focused material with special relevance to a single but important aspect of the Commission's program. Special Publications, being working documents, may evolve with the findings of and charges to a particular committee. Both publications follow the style of the Canadian Journal of Fisheries and Aquatic Sciences. Sponsorship of Technical Reports or Special Publications does not necessarily imply that the findings or conclusions contained therein are endorsed by the Commission.