5Ec33d32278a82020051901581
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Geological Survey of Ohio
GEOLOGICAL SURVEY OF OHIO. VOL. I.—PART II. PALÆONTOLOGY. SECTION II. DESCRIPTIONS OF FOSSIL FISHES. BY J. S. NEWBERRY. Digital version copyrighted ©2012 by Don Chesnut. THE CLASSIFICATION AND GEOLOGICAL DISTRIBUTION OF OUR FOSSIL FISHES. So little is generally known in regard to American fossil fishes, that I have thought the notes which I now give upon some of them would be more interesting and intelligible if those into whose hands they will fall could have a more comprehensive view of this branch of palæontology than they afford. I shall therefore preface the descriptions which follow with a few words on the geological distribution of our Palæozoic fishes, and on the relations which they sustain to fossil forms found in other countries, and to living fishes. This seems the more necessary, as no summary of what is known of our fossil fishes has ever been given, and the literature of the subject is so scattered through scientific journals and the proceedings of learned societies, as to be practically inaccessible to most of those who will be readers of this report. I. THE ZOOLOGICAL RELATIONS OF OUR FOSSIL FISHES. To the common observer, the class of Fishes seems to be well defined and quite distin ct from all the other groups o f vertebrate animals; but the comparative anatomist finds in certain unusual and aberrant forms peculiarities of structure which link the Fishes to the Invertebrates below and Amphibians above, in such a way as to render it difficult, if not impossible, to draw the lines sharply between these great groups. -

Chelicerata; Eurypterida) from the Campbellton Formation, New Brunswick, Canada Randall F
Document generated on 10/01/2021 9:05 a.m. Atlantic Geology Nineteenth century collections of Pterygotus anglicus Agassiz (Chelicerata; Eurypterida) from the Campbellton Formation, New Brunswick, Canada Randall F. Miller Volume 43, 2007 Article abstract The Devonian fauna from the Campbellton Formation of northern New URI: https://id.erudit.org/iderudit/ageo43art12 Brunswick was discovered in 1881 at the classic locality in Campbellton. About a decade later A.S. Woodward at the British Museum (Natural History) (now See table of contents the Natural History Museum, London) acquired specimens through fossil dealer R.F. Damon. Woodward was among the first to describe the fish assemblage of ostracoderms, arthrodires, acanthodians and chondrichthyans. Publisher(s) At the same time the museum also acquired specimens of a large pterygotid eurypterid. Although the vertebrates received considerable attention, the Atlantic Geoscience Society pterygotids at the Natural History Museum, London are described here for the first time. The first pterygotid specimens collected in 1881 by the Geological ISSN Survey of Canada were later identified by Clarke and Ruedemann in 1912 as Pterygotus atlanticus, although they suggested it might be a variant of 0843-5561 (print) Pterygotus anglicus Agassiz. An almost complete pterygotid recovered in 1994 1718-7885 (digital) from the Campbellton Formation at a new locality in Atholville, less than two kilometres west of Campbellton, has been identified as P. anglicus Agassiz. Like Explore this journal the specimens described by Clarke and Ruedemann, the material from the Natural History Museum, London is herein referred to P. anglicus. Cite this article Miller, R. F. (2007). Nineteenth century collections of Pterygotus anglicus Agassiz (Chelicerata; Eurypterida) from the Campbellton Formation, New Brunswick, Canada. -

Note on Pterygotus Anglicus Agassiz (Eurypterida: Devonian) from the Campbellton Formation, New Brunswick Randall F
Document generated on 10/01/2021 12:30 a.m. Atlantic Geology Note on Pterygotus anglicus Agassiz (Eurypterida: Devonian) from the Campbellton Formation, New Brunswick Randall F. Miller Volume 32, Number 2, Summer 1996 Article abstract Fragments of the large euryptcrid Pterygotus, recently collected from the URI: https://id.erudit.org/iderudit/ageo32_2art01 Devonian Campbellton Formation at Atholville, New Brunswick, are identified as belonging to P. anglicus Agassiz. The only previous Pterygotus specimens See table of contents from this site, collected in 1881, were assigned to a new species P. atlanticus Clarke and Rucdemann, in 1912. Clarke and Rucdcmann's suggestion that P. atlanticus might turn out to be a small specimen of P. anglicus is supported by Publisher(s) this new find. However, possible revision of P. atlanticus awaits the discovery of additional, more complete, material. Atlantic Geoscience Society ISSN 0843-5561 (print) 1718-7885 (digital) Explore this journal Cite this article Miller, R. F. (1996). Note on Pterygotus anglicus Agassiz (Eurypterida: Devonian) from the Campbellton Formation, New Brunswick. Atlantic Geology, 32(2), 95–100. All rights reserved © Atlantic Geology, 1996 This document is protected by copyright law. Use of the services of Érudit (including reproduction) is subject to its terms and conditions, which can be viewed online. https://apropos.erudit.org/en/users/policy-on-use/ This article is disseminated and preserved by Érudit. Érudit is a non-profit inter-university consortium of the Université de Montréal, Université Laval, and the Université du Québec à Montréal. Its mission is to promote and disseminate research. https://www.erudit.org/en/ A tlantic Geology 95 Note on Pterygotus anglicus Agassiz (Eurypterida: Devonian) from the Campbellton Formation, New Brunswick Randall F. -

Tayside, Central and Fife Tayside, Central and Fife
Detail of the Lower Devonian jawless, armoured fish Cephalaspis from Balruddery Den. © Perth Museum & Art Gallery, Perth & Kinross Council Review of Fossil Collections in Scotland Tayside, Central and Fife Tayside, Central and Fife Stirling Smith Art Gallery and Museum Perth Museum and Art Gallery (Culture Perth and Kinross) The McManus: Dundee’s Art Gallery and Museum (Leisure and Culture Dundee) Broughty Castle (Leisure and Culture Dundee) D’Arcy Thompson Zoology Museum and University Herbarium (University of Dundee Museum Collections) Montrose Museum (Angus Alive) Museums of the University of St Andrews Fife Collections Centre (Fife Cultural Trust) St Andrews Museum (Fife Cultural Trust) Kirkcaldy Galleries (Fife Cultural Trust) Falkirk Collections Centre (Falkirk Community Trust) 1 Stirling Smith Art Gallery and Museum Collection type: Independent Accreditation: 2016 Dumbarton Road, Stirling, FK8 2KR Contact: [email protected] Location of collections The Smith Art Gallery and Museum, formerly known as the Smith Institute, was established at the bequest of artist Thomas Stuart Smith (1815-1869) on land supplied by the Burgh of Stirling. The Institute opened in 1874. Fossils are housed onsite in one of several storerooms. Size of collections 700 fossils. Onsite records The CMS has recently been updated to Adlib (Axiel Collection); all fossils have a basic entry with additional details on MDA cards. Collection highlights 1. Fossils linked to Robert Kidston (1852-1924). 2. Silurian graptolite fossils linked to Professor Henry Alleyne Nicholson (1844-1899). 3. Dura Den fossils linked to Reverend John Anderson (1796-1864). Published information Traquair, R.H. (1900). XXXII.—Report on Fossil Fishes collected by the Geological Survey of Scotland in the Silurian Rocks of the South of Scotland. -

BMC Evolutionary Biology Biomed Central
BMC Evolutionary Biology BioMed Central Research article Open Access Evolution of miniaturization and the phylogenetic position of Paedocypris, comprising the world's smallest vertebrate Lukas Rüber*1, Maurice Kottelat2, Heok Hui Tan3, Peter KL Ng3 and Ralf Britz1 Address: 1Department of Zoology, The Natural History Museum, Cromwell Road, London SW7 5BD, UK, 2Route de la Baroche 12, Case postale 57, CH-2952 Cornol, Switzerland (permanent address) and Raffles Museum of Biodiversity Research, National University of Singapore, Kent Ridge, Singapore 119260 and 3Department of Biological Sciences, National University of Singapore, Kent Ridge, Singapore 119260 Email: Lukas Rüber* - [email protected]; Maurice Kottelat - [email protected]; Heok Hui Tan - [email protected]; Peter KL Ng - [email protected]; Ralf Britz - [email protected] * Corresponding author Published: 13 March 2007 Received: 23 October 2006 Accepted: 13 March 2007 BMC Evolutionary Biology 2007, 7:38 doi:10.1186/1471-2148-7-38 This article is available from: http://www.biomedcentral.com/1471-2148/7/38 © 2007 Rüber et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Abstract Background: Paedocypris, a highly developmentally truncated fish from peat swamp forests in Southeast Asia, comprises the world's smallest vertebrate. Although clearly a cyprinid fish, a hypothesis about its phylogenetic position among the subfamilies of this largest teleost family, with over 2400 species, does not exist. -

GY 112 Lecture Notes D
GY 112 lecture notes D. Haywick (2006) 1 GY 112 Lecture Notes Evolution of the Chordates Lecture Goals: A) The first animals with backbones (conodonts) B) The fish family tree C) Other key chordate evolutionary events Textbook reference: Levin 7th edition (2003) Chapter 10; Levin 8th edition (2006) Chapter 12 A) The conodonts By now you should realize that there are still a lot of mysteries in Earth history. The same can be said about biology, chemistry and physics. Science does not have all the answers, nor will it ever find all the answers. However, every once in a while, a long standing mystery or problem is solved. I bet you're thinking that in order to do this, a clever scientist had to dedicate his or her entire life to finding the solution. He/She had to work non- stop, day and night, taking a break only to eat, sleep for a few hours, and, occasionally, pee. Yep, sometimes this is what's done. Other times, however, it's just blind luck. Such is the case with a formerly mysterious group of animal remains simply classified as the conodonts. These hard body parts (see scanning electron microscope image to the left from, http://earthnet-geonet.ca/earth/ranges_e.php?s=conodonts) are usually quite small (less than 1 mm), are composed of the mineral apatite [Ca5(PO4)3(OH,F)] and really look like tiny teeth. They have been long used for biostratigraphy and there are many well-know examples of conodont index fossils, but apart from the fact that they look like teeth, and that they are now all extinct, no one really knew what they were. -

Phylum Chordata with Its Characters
Classification of phylum Chordata with its characters: TIG The chordates form a large heterogeneous group of members differing widely from one another in many respects. Due to great diversity in chordate forms, different schemes of classification have been proposed by a number of taxonomists from time to time. The classification followed is simplified and is a synthesis of most recent classifications. Table 1.2 gives an outline classification of the phylum Chordata. Phylum Chordata: Widely diversified (differing) in size, habits and habitat, bilaterally symmetrical, metamerically segmented, triploblastic, coelomate deuterostomes. All the chordates possess a supporting skeletal rod or notochord, a hollow dorsal nerve cord and paired gill-slits at some stage of their life history which may persist, change or disappear in adults. Cambrian to Recent. About 50,000 species. Phylum Chordata can be divided into two groups: A. Acrania (Protochordata) and B. Craniata (Euchordata) which show contrasting characters. Group A. Acrania (Protochordata): (Gr., a = absent; kranion = head or Gr., protos = first; chorde = cord). All are marine, small, primitive or lower chordates. No cranium, jaws, vertebral column, paired appendages. About 2,000 species. The Acrania is divided into three subphyla- Hemichordata, Urochordata and Cephalochordata. Subphylum I. Hemichordata: Gr., hemi = half; chorde = cord). Body divided into 3 regions- proboscis, collar and trunk. Notochord doubtful, short confined to proboscis and non-homologous with that of chordates. Class 1. Enteropneusta: (Gr., enteron = gut; pneustos = breathed). Body large and worm-like. Gill-slits numerous and paired. Alimentary canal straight. Acorn or tongue worms. Enteropneusts include 3 families, 15 genera and 70 species. Examples- Balanoglossus, Saccoglossus, Ptychodera. -
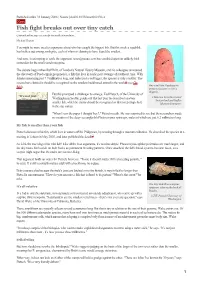
Fish Fight Breaks out Over Tiny Catch
Published online 31 January 2006 | Nature | doi:10.1038/news060130-4 News Fish fight breaks out over tiny catch Contenders line up to net credit for smallest vertebrate. Michael Hopkin You might be more used to arguments about who has caught the biggest fish. But this week a squabble has broken out among zoologists, each of whom is claiming to have found the smallest. And now, in attempting to settle the argument, [email protected] has stumbled upon an unlikely third contender for the small-vertebrate prize. The debate began when Ralf Britz, of London's Natural History Museum, and his colleagues announced the discovery of Paedocypris progenetica, a fish that lives in acidic peat swamps of southeast Asia. With females measuring just 7.9 millimetres long, and males just a tad bigger, the species is truly a tiddler. The researchers claimed it should be recognized as the smallest backboned animal in the world1 (see Go One small fish: Paedocypris fish!). progenetica gets lost on a fingertip. But this prompted a challenger to emerge. Ted Pietsch, of the University of “It's not just Washington in Seattle, points out that last year he described an even © Maurice Kottelat Cornol millimetres that Switzerland and Raffles count - it's how smaller fish, which he claims should be recognized as first (or perhaps last) Museum Singapore you use those in the size stakes. millimetres.” "When I saw the paper I thought 'hey!'," Pietsch recalls. He was surprised to see that the researchers made no mention of the deep-sea anglerfish Photocorynus spiniceps, males of which are just 6.2 millimetres long. -

Morphology and Histology of Acanthodian Fin Spines from the Late Silurian Ramsasa E Locality, Skane, Sweden Anna Jerve, Oskar Bremer, Sophie Sanchez, Per E
Morphology and histology of acanthodian fin spines from the late Silurian Ramsasa E locality, Skane, Sweden Anna Jerve, Oskar Bremer, Sophie Sanchez, Per E. Ahlberg To cite this version: Anna Jerve, Oskar Bremer, Sophie Sanchez, Per E. Ahlberg. Morphology and histology of acanthodian fin spines from the late Silurian Ramsasa E locality, Skane, Sweden. Palaeontologia Electronica, Coquina Press, 2017, 20 (3), pp.20.3.56A-1-20.3.56A-19. 10.26879/749. hal-02976007 HAL Id: hal-02976007 https://hal.archives-ouvertes.fr/hal-02976007 Submitted on 23 Oct 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Palaeontologia Electronica palaeo-electronica.org Morphology and histology of acanthodian fin spines from the late Silurian Ramsåsa E locality, Skåne, Sweden Anna Jerve, Oskar Bremer, Sophie Sanchez, and Per E. Ahlberg ABSTRACT Comparisons of acanthodians to extant gnathostomes are often hampered by the paucity of mineralized structures in their endoskeleton, which limits the potential pres- ervation of phylogenetically informative traits. Fin spines, mineralized dermal struc- tures that sit anterior to fins, are found on both stem- and crown-group gnathostomes, and represent an additional potential source of comparative data for studying acantho- dian relationships with the other groups of early gnathostomes. -

Dental Diversity in Early Chondrichthyans
1 Supplementary information 2 3 Dental diversity in early chondrichthyans 4 and the multiple origins of shedding teeth 5 6 Dearden and Giles 7 8 9 This PDF file includes: 10 Supplementary figures 1-5 11 Supplementary text 12 Supplementary references 13 Links to supplementary data 14 15 16 Supplementary Figure 1. Taemasacanthus erroli left lower jaw NHMUK PV 17 P33706 in (a) medial; (b) dorsal; (c) lateral; (d) ventral; (e) posterior; and (f) 18 dorsal and (g) dorso-medial views with tooth growth series coloured. Colours: 19 blue, gnathal plate; grey, Meckel’s cartilage. 20 21 Supplementary Figure 2. Atopacanthus sp. right lower or left upper gnathal 22 plate NHMUK PV P.10978 in (a) medial; (b) dorsal;, (c) lateral; (d) ventral; and 23 (e) dorso-medial view with tooth growth series coloured. Colours: blue, 24 gnathal plate. 25 26 Supplementary Figure 3. Ischnacanthus sp. left lower jaw NHMUK PV 27 P.40124 (a,b) in lateral view superimposed on digital mould of matrix surface 28 with Meckel’s cartilage removed in (b); (c) in lateral view; and (d) in medial 29 view. Colours: blue, gnathal plate; grey, Meckel’s cartilage. 30 Supplementary Figure 4. Acanthodopsis sp. right lower jaw NHMUK PV 31 P.10383 in (a,b) lateral view with (b) mandibular splint removed; (c) medial 32 view; (d) dorsal view; (e) antero-medial view, and (f) posterior view. Colours: 33 blue, teeth; grey, Meckel’s cartilage; green, mandibular splint. 34 35 Supplementary Figure 5. Acanthodes sp. Left and right lower jaws in 36 NHMUK PV P.8065 (a) viewed in the matrix, in dorsal view; (b) superimposed 37 on the digital mould of the matrix’s surface, in ventral view; and (c,d) the left 38 lower jaw isolated in (c) medial, and (d) lateral view. -

Evolutionary Trends of the Pharyngeal Dentition in Cypriniformes (Actinopterygii: Ostariophysi)
Evolutionary trends of the pharyngeal dentition in Cypriniformes (Actinopterygii: Ostariophysi). Emmanuel Pasco-Viel, Cyril Charles, Pascale Chevret, Marie Semon, Paul Tafforeau, Laurent Viriot, Vincent Laudet To cite this version: Emmanuel Pasco-Viel, Cyril Charles, Pascale Chevret, Marie Semon, Paul Tafforeau, et al.. Evolution- ary trends of the pharyngeal dentition in Cypriniformes (Actinopterygii: Ostariophysi).. PLoS ONE, Public Library of Science, 2010, 5 (6), pp.e11293. 10.1371/journal.pone.0011293. hal-00591939 HAL Id: hal-00591939 https://hal.archives-ouvertes.fr/hal-00591939 Submitted on 31 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Evolutionary Trends of the Pharyngeal Dentition in Cypriniformes (Actinopterygii: Ostariophysi) Emmanuel Pasco-Viel1, Cyril Charles3¤, Pascale Chevret2, Marie Semon2, Paul Tafforeau4, Laurent Viriot1,3*., Vincent Laudet2*. 1 Evo-devo of Vertebrate Dentition, Institut de Ge´nomique Fonctionnelle de Lyon, Universite´ de Lyon, CNRS, INRA, Ecole Normale Supe´rieure de Lyon, Lyon, France, 2 Molecular Zoology, Institut de Ge´nomique Fonctionnelle de Lyon, Universite´ de Lyon, CNRS, INRA, Ecole Normale Supe´rieure de Lyon, Lyon, France, 3 iPHEP, CNRS UMR 6046, Universite´ de Poitiers, Poitiers, France, 4 European Synchrotron Radiation Facility, Grenoble, France Abstract Background: The fish order Cypriniformes is one of the most diverse ray-finned fish groups in the world with more than 3000 recognized species. -

Ing.Org Published in Great Britain by the Royal Society, 6–9 Carlton House Terrace, London SW1Y 5AG Coturnix Coturnix Japonica Number 1665 Number
RSPB_276_1665_Cover.qxp 4/23/09 4:39 PM Page 1 Proc. R. Soc. B | vol. 276 no. 1665 pp. 2133–2331 22 June 2009 ISSN 0962-8452 volume 276 22 June 2009 number 1665 volume 276 . number 1665 . pages 2133–2331 pages 2133–2331 Review articles Chloroplast two-component systems: evolution of the link between photosynthesis and gene expression 2133 S. Puthiyaveetil & J. F. Allen Research articles Going to great lengths: selection for long corolla tubes in an extremely specialized bat–flower mutualism 2147 N. Muchhala & J. D. Thomson Dynamics of crowing development in the domestic Japanese quail (Coturnix coturnix japonica) 2153 S. Derégnaucourt, S. Saar & M. Gahr Emperor penguin mates: keeping together in the crowd 2163 A. Ancel, M. Beaulieu, Y. Le Maho & C. Gilbert A quantum probability explanation for violations of ‘rational’ decision theory 2171 E. M. Pothos & J. R. Busemeyer Spectacular morphological novelty in a miniature cyprinid fish, Danionella dracula n. sp. 2179 R. Britz, K. W. Conway & L. Rüber Does colour polymorphism enhance survival of prey populations? 2187 L. Wennersten & A. Forsman Genetic linkage map of the guppy, Poecilia reticulata, and quantitative trait loci analysis of male size and colour variation 2195 N. Tripathi, M. Hoffmann, E.-M. Willing, C. Lanz, D. Weigel & C. Dreyer Biodiversity and body size are linked across metazoans 2209 C. R. McClain & A. G. Boyer The evolution of covert, silent infection as a parasite strategy 2217 I. Sorrell, A. White, A. B. Pedersen, R. S. Hails & M. Boots Avian orientation: the pulse effect is mediated by the magnetite receptors in the upper beak 2227 W.