Preliminary Post-Cranial Metric Analysis of Mammoths from the Hot Springs Mammoth Site, South Dakota
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pleistocene Mammals from Extinction Cave, Belize
Canadian Journal of Earth Sciences Pleistocene Mammals From Extinction Cave, Belize Journal: Canadian Journal of Earth Sciences Manuscript ID cjes-2018-0178.R3 Manuscript Type: Article Date Submitted by the 04-May-2019 Author: Complete List of Authors: Churcher, C.S.; University of Toronto, Zoology Central America, Pleistocene, Fauna, Vertebrate Palaeontology, Keyword: Limestone cave Is the invited manuscript for consideration in a Special Not applicableDraft (regular submission) Issue? : https://mc06.manuscriptcentral.com/cjes-pubs Page 1 of 43 Canadian Journal of Earth Sciences 1 1 PLEISTOCENE MAMMALS FROM EXTINCTION CAVE, BELIZE 2 by C.S. CHURCHER1 Draft 1Department of Zoology, University of Toronto, Toronto, Ontario Canada M5S 2C6 and 322-240 Dallas Rd., Victoria, British Columbia, Canada V8V 4X9 (corresponding address): e-mail [email protected] https://mc06.manuscriptcentral.com/cjes-pubs Canadian Journal of Earth Sciences Page 2 of 43 2 4 5 ABSTRACT. A small mammalian fauna is recorded from Extinction Cave (also called Sibun 6 Cave), east of Belmopan, on the Sibun River, Belize, Central America. The animals recognized 7 are armadillo (†Dasypus bellus), American lion (†Panthera atrox), jaguar (P. onca), puma or 8 mountain lion (Puma concolor), Florida spectacled bear (†Tremarctos floridanus), javelina or 9 collared peccary (Pecari tajacu), llama (Camelidae indet., ?†Palaeolama mirifica), red brocket 10 deer (Mazama americana), bison (Bison sp.) and Mexican half-ass (†Equus conversidens), and 11 sabre-tooth cat († Smilodon fatalis) may also be represented (‘†’ indicates an extinct taxon). 12 Bear and bison are absent from the region today. The bison record is one of the more southernly 13 known. The bear record is almost the mostDraft westerly known and a first for Central America. -

Palaeontologia Electronica COMPARATIVE VARIABILITY OF
Palaeontologia Electronica http://palaeo-electronica.org COMPARATIVE VARIABILITY OF INTERMEMBRANOUS AND ENDOCHONDRAL BONES IN PLEISTOCENE MAMMALS Kristina R. Raymond and Donald R. Prothero ABSTRACT Study of the embryology and ossification of modern bones predicts that fossil intermembranous bones (which ossify from connective tissue) will exhibit greater size variability than endochondral bones (which are formed from embryological cartilagi- nous precursors), because intermembranous bones are less tightly constrained by joints and articular surfaces. To evaluate this hypothesis, we measured multiple dimen- sions of 989 intermembranous bones (patellae and other sesamoids) of the saber- toothed cat Smilodon fatalis, the Ice Age lion Panthera atrox, the bison Bison antiquus, the horse Equus occidentalis, the camel Camelops hesternus, the ground sloths Paramylodon (=Glossotherium) harlani and Nothrotheriops shastensis from Rancho La Brea and from the late Pleistocene San Josecito Cave in Nuevo Leon, Mexico. These were compared to measurements of 811 endochondral bones (primarily astragali) of comparable size. Through statistical analyses (coefficients of variation, ANOVA, modi- fied Levene’s test, and t-tests) we found slight evidence of higher variability in many of the intermembranous bones of these taxa (21 out of 27 CVs were higher for intermem- branous bones than endochondral bones), although this trend is not found in all taxa. Using a modified Levene’s test, only Smilodon and some of the dimensions of horse and bison patellae are significantly more variable than the corresponding dimensions of the astragali. Although the results are mixed, at least some data show that inter- membranous bones are not as tightly constrained by growth and by adjacent tissues as are endochondral bones. -

Prospects for Rewilding with Camelids
Journal of Arid Environments 130 (2016) 54e61 Contents lists available at ScienceDirect Journal of Arid Environments journal homepage: www.elsevier.com/locate/jaridenv Prospects for rewilding with camelids Meredith Root-Bernstein a, b, *, Jens-Christian Svenning a a Section for Ecoinformatics & Biodiversity, Department of Bioscience, Aarhus University, Aarhus, Denmark b Institute for Ecology and Biodiversity, Santiago, Chile article info abstract Article history: The wild camelids wild Bactrian camel (Camelus ferus), guanaco (Lama guanicoe), and vicuna~ (Vicugna Received 12 August 2015 vicugna) as well as their domestic relatives llama (Lama glama), alpaca (Vicugna pacos), dromedary Received in revised form (Camelus dromedarius) and domestic Bactrian camel (Camelus bactrianus) may be good candidates for 20 November 2015 rewilding, either as proxy species for extinct camelids or other herbivores, or as reintroductions to their Accepted 23 March 2016 former ranges. Camels were among the first species recommended for Pleistocene rewilding. Camelids have been abundant and widely distributed since the mid-Cenozoic and were among the first species recommended for Pleistocene rewilding. They show a range of adaptations to dry and marginal habitats, keywords: Camelids and have been found in deserts, grasslands and savannas throughout paleohistory. Camelids have also Camel developed close relationships with pastoralist and farming cultures wherever they occur. We review the Guanaco evolutionary and paleoecological history of extinct and extant camelids, and then discuss their potential Llama ecological roles within rewilding projects for deserts, grasslands and savannas. The functional ecosystem Rewilding ecology of camelids has not been well researched, and we highlight functions that camelids are likely to Vicuna~ have, but which require further study. -

Paleobiology of a Large Mammal Community from the Late Pleistocene of Sonora, Mexico
Quaternary Research (2021), 102, 247–259 doi:10.1017/qua.2020.125 Research Article Paleobiology of a large mammal community from the late Pleistocene of Sonora, Mexico Rachel A. Shorta* , Laura G. Emmertb, Nicholas A. Famosoc,d, Jeff M. Martina,†, Jim I. Meade,f, Sandy L. Swifte and Arturo Baezg aDepartment of Ecology and Conservation Biology, Texas A&M University, College Station, Texas 77843 USA; bDon Sundquist Center of Excellence in Paleontology, East Tennessee State University, Johnson City, Tennessee 37614 USA; cJohn Day Fossil Beds National Monument, U.S. National Park Service, Kimberly, Oregon 97848 USA; dDepartment of Earth Sciences, University of Oregon, Eugene, Oregon 97403 USA; eThe Mammoth Site of Hot Springs, S.D.,1800 Hwy 18 Bypass, Hot Springs, South Dakota, 57747 USA; fDesert Laboratory on Tumamoc Hill, University of Arizona, Tucson, Arizona 85745 USA and gCollege of Agriculture and Life Sciences, University of Arizona, Tucson, Arizona 85721 USA Abstract A paleontological deposit near San Clemente de Térapa represents one of the very few Rancholabrean North American Land Mammal Age sites within Sonora, Mexico. During that time, grasslands were common, and the climate included cooler and drier summers and wetter winters than currently experienced in northern Mexico. Here, we demonstrate restructuring in the mammalian community associated with environmental change over the past 40,000 years at Térapa. The fossil community has a similar number of carnivores and herbivores whereas the modern community consists mostly of carnivores. There was also a 97% decrease in mean body size (from 289 kg to 9 kg) because of the loss of megafauna. -

Teacher's Guide
Waco Mammoth National Monument Teacher’s Guide Waco Mammoth National Monument is a proud partnership: COVID-19 Safety Notice Wearing a mask is required in federal buildings. The Dig Shelter building at Waco Mammoth National Monument has a maximum capacity of 25 individuals. This document updated 5 May 2021. 1 Park Information Address: 6220 Steinbeck Bend Drive Waco, Texas 76708 Front Office: 254-299-2663 Unload buses at the and then use the BUS PARKING. Bathrooms are in the . The walk from the to the DIG SHELTER is 300 yards. No food or drink is allowed in the DIG SHELTER. Map data ©Google, 2021. All photos and illustrations in this guide, unless otherwise stated, have been provided by the City of Waco. 2 I. Dig Shelter Guide Begin at the entrance bridge, shown in the image below. The following pages provide a helpful script as well as critical thinking questions for your students. 3 A. History of the Site All the fossils you will see today are from the Pleistocene Epoch, or the Ice Age. The Ice Age started 2.6 million years ago and ended just 12,000 years ago. SAY IT TOGETHER: PLY-stuh-SEEN EP-uck This bridge crosses over the first dig site. In 1978, two young men named Paul Barron and Eddie Bufkin came to explore this creek, hoping to find fossils, or possibly some arrowheads. They found a very large bone near the embankment you see by the bridge. They didn’t know what the bone was, so they took it to Baylor University. -

Arms Race of Temporal Partitioning Between Carnivorous And
www.nature.com/scientificreports OPEN Arms race of temporal partitioning between carnivorous and herbivorous mammals Received: 26 October 2017 Yonghua Wu1,2, Haifeng Wang3, Haitao Wang4 & Jiang Feng2,5 Accepted: 12 January 2018 Reciprocal coevolutionary changes in predation and anti-predator behaviours have long been Published: xx xx xxxx hypothesized, but evolutionary-scale evidence is rare. Here, we reconstructed the evolutionary-scale changes in the diel activity patterns of a predator-prey system (carnivorous and herbivorous mammals) based on a molecular phyloecological approach, providing evidence of long-term antagonistic coevolutionary changes in their diel activities. Our molecular reconstruction of diel activity patterns, which is supported by morphological evidence, consistently showed that carnivorous mammals were subjected to a shift from diurnality to nocturnality, while herbivorous mammals experienced a shift from nocturnality to diurnality during their evolutionary histories. A shift in the diel activity of the herbivores as a result of carnivore avoidance is hypothesized based on molecular, morphological and behavioural evidence, and our results suggest an evolutionary-scale arms race of diel activity shifts between carnivorous and herbivorous mammals. Interactions between carnivorous and herbivorous mammals, representing one of the classic coevolutionary sys- tems, lead to long-term reciprocal evolutionary changes in predation and anti-predator behaviours1. Among carnivorous mammals, felids (Felidae) and canids (Canidae) are the main predators of herbivorous mammals (e.g., ungulates)2. Tese carnivores (felids and canids) and ungulates show diferentiated diel activity patterns, with most felids and canids being mainly nocturnal, while ungulates are primarily diurnal3,4. Given the difer- entiation of their diel activity patterns, one possibility is that the diurnality of ungulates may have evolved as an anti-predator behaviour. -

Giant Camels from the Cenozoic of North America SERIES PUBLICATIONS of the SMITHSONIAN INSTITUTION
Giant Camels from the Cenozoic of North America SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the Institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Contributions to Anthropology Smithsonian Contributions to Astrophysics Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to the H^arine Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoology Smithsonian Folklife Studies Smithsonian Studies in Air and Space Smithsonian Studies in History and Technology In these series, the Institution publishes small papers and full-scale monographs that refXJrt the research and collections of its various museums and bureaux or of professional colleagues in the world of science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the worid. Papers or monographs submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. Press requirements for manuscript and art preparation are outlined on the inside back cover. -

Ancient Bison (Bison Antiquus)
Ancient bison (Bison antiquus) The ancient bison grazed in the valleys eating grasses, shrubs and woody plants. Ancient bison became extinct about 10,000 years ago. The ancient bison is the ancestor of today’s bison. Ancient bison traveled in herds or groups. What is the name of another animal that traveled in herds? 2345 Searl Parkway, Hemet, CA 92543 www.WesternScienceCenter.org “Yesterday’s” camel (Camelops hesternus) The “Yesterday’s” camel is very closely related to today’s living Ilama. Camels originally lived in North America 50 million years ago before spreading to other parts of the world. “Yesterday’s” camel is now extinct. Camels are herbivores and eat lots of grasses. Unlike camels that live today “Yesterday’s” camel may have liked to eat leafy forest plants. Why do scientists with the Diamond Valley Lake project think “Yesterday’s” camel lived in herds or large groups? 2345 Searl Parkway, Hemet, CA 92543 www.WesternScienceCenter.org Dire wolf (Canis dirus) Fossils of the dire wolf were found while digging the Diamond Valley Lake project. This extinct wolf was as big as the gray wolf that lives today. The jaws and large teeth of the dire wolf were the most powerful of all the wolves. This would help them catch their food. The dire wolf became extinct about 11,000 years ago. The word "dire" means “grim.” Why do you think this wolf was named "dire" wolf? 2345 Searl Parkway, Hemet, CA 92543 www.WesternScienceCenter.org Western horse (Equus occidentalis) The Western horse lived in large herds or groups. They were as tall as today’s Arabian horse. -
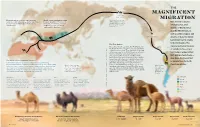
Magnificent Migration
THE MAGNIFICENT Land bridge 8 MY–14,500 Y MIGRATION Domestication of camels began between World camel population today About 6 million years ago, 3,000 and 4,000 years ago—slightly later than is about 30 million: 27 million of camelids began to move Best known today for horses—in both the Arabian Peninsula and these are dromedaries; 3 million westward across the land Camelini western Asia. are Bactrians; and only about that connected Asia and inhabiting hot, arid North America. 1,000 are Wild Bactrians. regions of North Africa and the Middle East, as well as colder steppes and Camelid ancestors deserts of Asia, the family Camelidae had its origins in North America. The The First Camels signature physical features The earliest-known camelids, the Protylopus and Lamini E the Poebrotherium, ranged in sizes comparable to of camels today—one or modern hares to goats. They appeared roughly 40 DMOR I SK million years ago in the North American savannah. two humps, wide padded U L Over the 20 million years that followed, more U L ; feet, well-protected eyes— O than a dozen other ancestral members of the T T O family Camelidae grew, developing larger bodies, . E may have developed K longer legs and long necks to better browse high C I The World's Most Adaptable Traveler? R ; vegetation. Some, like Megacamelus, grew even I as adaptations to North Camels have adapted to some of the Earth’s most demanding taller than the woolly mammoths in their time. LEWSK (Later, in the Middle East, the Syrian camel may American winters. -

Medial Pleistocene Fossil Vertebrate Localities in Nebraska
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Bulletin of the University of Nebraska State Museum Museum, University of Nebraska State 1957 Medial Pleistocene Fossil Vertebrate Localities in Nebraska C. Bertrand Schultz Lloyd G. Tanner Follow this and additional works at: https://digitalcommons.unl.edu/museumbulletin Part of the Entomology Commons, Geology Commons, Geomorphology Commons, Other Ecology and Evolutionary Biology Commons, Paleobiology Commons, Paleontology Commons, and the Sedimentology Commons Schultz, C. Bertrand and Tanner, Lloyd G., "Medial Pleistocene Fossil Vertebrate Localities in Nebraska" (1957). Bulletin of the University of Nebraska State Museum. 14. https://digitalcommons.unl.edu/museumbulletin/14 This Article is brought to you for free and open access by the Museum, University of Nebraska State at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Bulletin of the University of Nebraska State Museum by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. BULLETIN OF' The University of Nebraska State Museum VOLUME 4, NUMBER 4 SEPTEMBER 1957 c. Bertrand Schultz Lloyd G. Tanner Medial Pleistocene Fossil Vertebrate Localities in Nebraska UNIVERSITY OF NEBRASKA Clifford M Hardin, Chancellor Board of Regents J. G. ELLIOTT CLARENCE E. SWANSON FRANK FOOTE CHARLES Y. THOMPSON B. N. GREENBERG, M.D. J. LEROY WELSH JOHN KENT SELLECK, Corporation Secretary Museum Staff C. BERTRAND SCHULTZ, Director; NATHAN L. MOHLER, Staff Curator, Vertebrate Artist Paleontology RALPH S. MUELLER, JACK BRODIE, Artist Honorary Curator, JOHN F. DAVIDSON, Curator, Health Sciences Botany and Herbarium LARRY W. QUATE, Curator, AUGUST EICHE, Honorary Entomology Curator, Ornithology HENRY REIDER, Chief FRANKLIN FENENGA, Associate Preparator Curator, Anthropology JOHN M. -

Paleontological Resource Inventory and Monitoring, Upper Columbia Basin Network
National Park Service U.S. Department of the Interior Upper Columbia Basin Network Paleontological Resource Inventory and Monitoring UPPER COLUMBIA BASIN NETWORK Paleontological Resource Inventory and Monitoring \ UPPER COLUMBIA BASIN NETWORK Jason P. Kenworthy Inventory and Monitoring Contractor George Washington Memorial Parkway Vincent L. Santucci Chief Ranger George Washington Memorial Parkway Michaleen McNerney Paleontological Intern Seattle, WA Kathryn Snell Paleontological Intern Seattle, WA August 2005 National Park Service, TIC #D-259 NOTE: This report provides baseline paleontological resource data to National Park Service administration and resource management staff. The report contains information regarding the location of non-renewable paleontological resources within NPS units. It is not intended for distribution to the general public. On the Cover: Well-preserved skull of the “Hagerman Horse”, Equus simplicidens , from Hagerman Fossil Beds National Monument. Equus simplicidens is the earliest, most primitive known representative of the modern horse genus Equus and the state fossil of Idaho. For more information, see page 17. Photo: NPS/Smithsonian Institution. How to cite this document: Kenworthy, J.P., V. L. Santucci, M. McNerney, and K. Snell. 2005. Paleontological Resource Inventory and Monitoring, Upper Columbia Basin Network. National Park Service TIC# D-259. TABLE OF CONTENTS INTRODUCTION ...................................................................................................................................1 -

Selections from Papers of the MASAL--Vol. 43 – Page 1 of 10
SUMMARY OF NORTH AMERICAN summaries of the Pleistocene fossil record in North America. A brief discussion of the problems PLEISTOCENE MAMMALIAN LOCAL FAUNAS encountered in age assignment of local faunas and use of provincial age and stage terms as applied to the CLAUDE W. HIBBARD Pleistocene follows. An explanation is given of the University of Michigan stratigraphic assignment of the local faunas in Table I. PAPERS OF THE MICHIGAN ACADEMY OP SCIENCE, ARTS, AND LETTERS VOL. XLIII, 1958 (1957 MEETING) Printed in U.S.A. DIFFERENT CONCEPTS OF THE PLEISTOCENE INTRODUCTION Zeuner (1950, p. 126) wrote, “the concept of Pleistocene THE ages of various Pleistocene faunas have plagued time has developed along three almost independent geologists since the first recognition of glacial and lines.” I list and briefly discuss these three different interglacial intervals. The chief handicap in assigning concepts here, since few students outside of the field of Pleistocene faunas to different ages is the lack of faunas vertebrate paleontology are aware of these problems. from the glaciated regions of North America which could Typology—Lyell (1833) named certain units of Cenozoic provide controls for dating faunas from the nonglaciated rocks Older Pliocene and Newer Pliocene. He later regions. Faunas are unknown from tills, and only a few (1839) applied the name Pliocene to what he previously fossils are known from outwash gravel and sand or from called “Older Pliocene” and the name Pleistocene to the interglacial deposits in the glaciated regions. With the “Newer Pliocene.” Calabrian and Villafranchian exception of post-Sangamon deposits, no vertebrate fossiliferous beds are present in the type section of fauna represented by more than a few specimens has Lyell’s Pliocene.