An Overview of Species Concept Used in Spiders Taxonomy with an Emphasis on Mating Structures
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Summary List of Fossil Spiders
A summary list of fossil spiders compiled by Jason A. Dunlop (Berlin), David Penney (Manchester) & Denise Jekel (Berlin) Suggested citation: Dunlop, J. A., Penney, D. & Jekel, D. 2010. A summary list of fossil spiders. In Platnick, N. I. (ed.) The world spider catalog, version 10.5. American Museum of Natural History, online at http://research.amnh.org/entomology/spiders/catalog/index.html Last udated: 10.12.2009 INTRODUCTION Fossil spiders have not been fully cataloged since Bonnet’s Bibliographia Araneorum and are not included in the current Catalog. Since Bonnet’s time there has been considerable progress in our understanding of the spider fossil record and numerous new taxa have been described. As part of a larger project to catalog the diversity of fossil arachnids and their relatives, our aim here is to offer a summary list of the known fossil spiders in their current systematic position; as a first step towards the eventual goal of combining fossil and Recent data within a single arachnological resource. To integrate our data as smoothly as possible with standards used for living spiders, our list follows the names and sequence of families adopted in the Catalog. For this reason some of the family groupings proposed in Wunderlich’s (2004, 2008) monographs of amber and copal spiders are not reflected here, and we encourage the reader to consult these studies for details and alternative opinions. Extinct families have been inserted in the position which we hope best reflects their probable affinities. Genus and species names were compiled from established lists and cross-referenced against the primary literature. -

Monitoring Report Spring/Summer 2015 Contents
Wimbledon and Putney Commons Monitoring Report Spring/Summer 2015 Contents CONTEXT 1 A. SYSTEMATIC RECORDING 3 METHODS 3 OUTCOMES 6 REFLECTIONS AND RECOMMENDATIONS 18 B. BIOBLITZ 19 REFLECTIONS AND LESSONS LEARNT 21 C. REFERENCES 22 LIST OF FIGURES Figure 1 Location of The Plain on Wimbledon and Putney Commons 2 Figure 2 Experimental Reptile Refuge near the Junction of Centre Path and Somerset Ride 5 Figure 3 Contrasting Cut and Uncut Areas in the Conservation Zone of The Plain, Spring 2015 6/7 Figure 4 Notable Plant Species Recorded on The Plain, Summer 2015 8 Figure 5 Meadow Brown and white Admiral Butterflies 14 Figure 6 Hairy Dragonfly and Willow Emerald Damselfly 14 Figure 7 The BioBlitz Route 15 Figure 8 Vestal and European Corn-borer moths 16 LIST OF TABLES Table 1 Mowing Dates for the Conservation Area of The Plain 3 Table 2 Dates for General Observational Records of The Plain, 2015 10 Table 3 Birds of The Plain, Spring - Summer 2015 11 Table 4 Summary of Insect Recording in 2015 12/13 Table 5 Rare Beetles Living in the Vicinity of The Plain 15 LIST OF APPENDICES A1 The Wildlife and Conservation Forum and Volunteer Recorders 23 A2 Sward Height Data Spring 2015 24 A3 Floral Records for The Plain : Wimbledon and Putney Commons 2015 26 A4 The Plain Spring and Summer 2015 – John Weir’s General Reports 30 A5 a Birds on The Plain March to September 2015; 41 B Birds on The Plain - summary of frequencies 42 A6 ai Butterflies on The Plain (DW) 43 aii Butterfly long-term transect including The Plain (SR) 44 aiii New woodland butterfly transect -

Biodiversity of the Huautla Cave System, Oaxaca, Mexico
diversity Communication Biodiversity of the Huautla Cave System, Oaxaca, Mexico Oscar F. Francke, Rodrigo Monjaraz-Ruedas † and Jesús A. Cruz-López *,‡ Colección Nacional De Arácnidos, Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacán, Mexico City C. P. 04510, Mexico; [email protected] (O.F.F.); [email protected] (R.M.-R.) * Correspondence: [email protected] † Current address: San Diego State University, San Diego, CA 92182, USA. ‡ Current address: Instituto Nacional de Investigaciones Agrícolas y Pecuarias del Valle de Oaxaca, Santo Domingo Barrio Bajo, Etla C. P. 68200, Mexico. Abstract: Sistema Huautla is the deepest cave system in the Americas at 1560 m and the fifth longest in Mexico at 89,000 m, and it is a mostly vertical network of interconnected passages. The surface landscape is rugged, ranging from 3500 to 2500 masl, intersected by streams and deep gorges. There are numerous dolinas, from hundreds to tens of meters in width and depth. The weather is basically temperate subhumid with summer rains. The average yearly rainfall is approximately 2500 mm, with a monthly average of 35 mm for the driest times of the year and up to 500 mm for the wettest month. All these conditions play an important role for achieving the highest terrestrial troglobite diversity in Mexico, containing a total of 35 species, of which 27 are possible troglobites (16 described), including numerous arachnids, millipedes, springtails, silverfish, and a single described species of beetles. With those numbers, Sistema Huautla is one of the richest cave systems in the world. Keywords: troglobitics; arachnids; insects; millipedes Citation: Francke, O.F.; Monjaraz-Ruedas, R.; Cruz-López, J.A. -

Desktop Biodiversity Report
Desktop Biodiversity Report Land at Balcombe Parish ESD/14/747 Prepared for Katherine Daniel (Balcombe Parish Council) 13th February 2014 This report is not to be passed on to third parties without prior permission of the Sussex Biodiversity Record Centre. Please be aware that printing maps from this report requires an appropriate OS licence. Sussex Biodiversity Record Centre report regarding land at Balcombe Parish 13/02/2014 Prepared for Katherine Daniel Balcombe Parish Council ESD/14/74 The following information is included in this report: Maps Sussex Protected Species Register Sussex Bat Inventory Sussex Bird Inventory UK BAP Species Inventory Sussex Rare Species Inventory Sussex Invasive Alien Species Full Species List Environmental Survey Directory SNCI M12 - Sedgy & Scott's Gills; M22 - Balcombe Lake & associated woodlands; M35 - Balcombe Marsh; M39 - Balcombe Estate Rocks; M40 - Ardingly Reservior & Loder Valley Nature Reserve; M42 - Rowhill & Station Pastures. SSSI Worth Forest. Other Designations/Ownership Area of Outstanding Natural Beauty; Environmental Stewardship Agreement; Local Nature Reserve; National Trust Property. Habitats Ancient tree; Ancient woodland; Ghyll woodland; Lowland calcareous grassland; Lowland fen; Lowland heathland; Traditional orchard. Important information regarding this report It must not be assumed that this report contains the definitive species information for the site concerned. The species data held by the Sussex Biodiversity Record Centre (SxBRC) is collated from the biological recording community in Sussex. However, there are many areas of Sussex where the records held are limited, either spatially or taxonomically. A desktop biodiversity report from SxBRC will give the user a clear indication of what biological recording has taken place within the area of their enquiry. -

Section IV – Guideline for the Texas Priority Species List
Section IV – Guideline for the Texas Priority Species List Associated Tables The Texas Priority Species List……………..733 Introduction For many years the management and conservation of wildlife species has focused on the individual animal or population of interest. Many times, directing research and conservation plans toward individual species also benefits incidental species; sometimes entire ecosystems. Unfortunately, there are times when highly focused research and conservation of particular species can also harm peripheral species and their habitats. Management that is focused on entire habitats or communities would decrease the possibility of harming those incidental species or their habitats. A holistic management approach would potentially allow species within a community to take care of themselves (Savory 1988); however, the study of particular species of concern is still necessary due to the smaller scale at which individuals are studied. Until we understand all of the parts that make up the whole can we then focus more on the habitat management approach to conservation. Species Conservation In terms of species diversity, Texas is considered the second most diverse state in the Union. Texas has the highest number of bird and reptile taxon and is second in number of plants and mammals in the United States (NatureServe 2002). There have been over 600 species of bird that have been identified within the borders of Texas and 184 known species of mammal, including marine species that inhabit Texas’ coastal waters (Schmidly 2004). It is estimated that approximately 29,000 species of insect in Texas take up residence in every conceivable habitat, including rocky outcroppings, pitcher plant bogs, and on individual species of plants (Riley in publication). -

Stillingfleet Nurseries to Enable the Best of the Catches to Be Shown to the Visitors on Sunday 28 June
Moth trapping at Stillingfleet Lodge Nurseries (SE 589409) from June 2012 Onwards Following a request from Mrs Vanessa Cook for representatives of Butterfly Conservation, Yorkshire Branch to attend a “Conservation Day” in the grounds of Stillingfleet Lodge it was agreed to install moth traps within the grounds and to display the catches to the visiting public at the event. As with much of the early summer in 2012 the weather was not very conducive to flying insects, although a reasonable number of specimens were caught and retained in small see- through containers to enable them to be viewed by the visitors. In spite of the damp conditions a good number of visitors did attend to support the stall set up by Butterfly Conservation, alongside several other attendee conservation groups. The following reports are written by David Baker who was accompanied on all the visits, unless noted otherwise, by David laughlin. The word “trap” alarms some people who associate the word with killing etc. However, no moths were harmed in the exercise and all were released after counting and, in this instance, displaying to other interested parties during the afternoon. At the end of the day we agreed to carry out further trapping later on in the year and, perhaps in following years, to try to assess the species of moths flying in the area of the Lodge and the following lists show the results of further trapping at the site. The three 2012 sessions, separated by several weeks, showed a great variety of moths with repeat species being very few and a total of 94 species were recorded. -
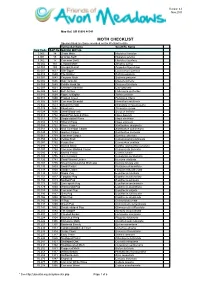
MOTH CHECKLIST Species Listed Are Those Recorded on the Wetland to Date
Version 4.0 Nov 2015 Map Ref: SO 95086 46541 MOTH CHECKLIST Species listed are those recorded on the Wetland to date. Vernacular Name Scientific Name New Code B&F No. MACRO MOTHS 3.005 14 Ghost Moth Hepialus humulae 3.001 15 Orange Swift Hepialus sylvina 3.002 17 Common Swift Hepialus lupulinus 50.002 161 Leopard Moth Zeuzera pyrina 54.008 169 Six-spot Burnet Zygaeba filipendulae 66.007 1637 Oak Eggar Lasiocampa quercus 66.010 1640 The Drinker Euthrix potatoria 68.001 1643 Emperor Moth Saturnia pavonia 65.002 1646 Oak Hook-tip Drepana binaria 65.005 1648 Pebble Hook-tip Drepana falcataria 65.007 1651 Chinese Character Cilix glaucata 65.009 1653 Buff Arches Habrosyne pyritoides 65.010 1654 Figure of Eighty Tethia ocularis 65.015 1660 Frosted Green Polyploca ridens 70.305 1669 Common Emerald Hermithea aestivaria 70.302 1673 Small Emerald Hemistola chrysoprasaria 70.029 1682 Blood-vein Timandra comae 70.024 1690 Small Blood-vein Scopula imitaria 70.013 1702 Small Fan-footed Wave Idaea biselata 70.011 1708 Single-dotted Wave Idaea dimidiata 70.016 1713 Riband Wave Idaea aversata 70.053 1722 Flame Carpet Xanthorhoe designata 70.051 1724 Red Twin-spot Carpet Xanthorhoe spadicearia 70.049 1728 Garden Carpet Xanthorhoe fluctuata 70.061 1738 Common Carpet Epirrhoe alternata 70.059 1742 Yellow Shell Camptogramma bilineata 70.087 1752 Purple Bar Cosmorhoe ocellata 70.093 1758 Barred Straw Eulithis (Gandaritis) pyraliata 70.097 1764 Common Marbled Carpet Chloroclysta truncata 70.085 1765 Barred Yellow Cidaria fulvata 70.100 1776 Green Carpet Colostygia pectinataria 70.126 1781 Small Waved Umber Horisme vitalbata 70.107 1795 November/Autumnal Moth agg Epirrita dilutata agg. -

IBEITR.ARANEOL.,L(2004)
I BEITR.ARANEOL.,l(2004) I PART 111 a (TEil 111 a) - Descriptions of selected taxa THE FOSSil MYGAlOMORPH SPIDERS (ARANEAE) IN BAl TIC AND DOMINICAN AMBER AND ABOUT EXTANT MEMBERS OF THE FAMllY MICROMYGALIDAE J. WUNDERLICH, 75334 Straubenhardt, Germany. Abstract: The fossil mygalomorph spiders (Araneae: Mygalomorpha) in Baltic and Do- minican amber are listed, a key to the taxa is given. Two species of the genus Ummidia THORELL 1875 (Ctenizidae: Pachylomerinae) in Baltic amber are redescribed, Clos- thes priscus MENGE 1869 (Dipluridae) from Baltic amber is revised, two gen. indet. (Dipluridae) fram Baltic amber are reported. The first fossil member of the family Micro- stigmatidae: Parvomygale n. gen., Parvomygale distineta n. sp. (Parvomygalinae n. subfarn.) in Dominican amber is described. - The taxon Micramygalinae PLATNICK & FORSTER 1982 is raised to family rank, revised diagnoses of the families Micromyga- lidae (no fossil record) and Micrastigmatidae are given. Material: CJW = collection J. WUNDERLICH, GPIUH = Geological and Palaeontologi- cal Institute of the University Hamburg, IMGPUG = Institute and Museum for Geology and Paleontology of the Georg-August-University Goettingen in Germany. 595 ---~-~-~~~--~~--~-'----------~--------~-~~~=-~~--.., INTRODUCTION The first fossil member of the suborder Mygalomorpha (= Orthognatha) in Baltic amber has been described by MENGE 1869 as Glostes priscus (figs. 1-2; comp. the book of WUNDERLICH (1986: Fig. 291)). This spider is a member of the family Dipluridae (Funnelweb Mygalomorphs) and is redescribed in this paper; only juveniles are known. Two further species of Mygalomorpha are described from this kind of amber, these are members of the family Ctenizidae (Trapdoor spiders). - Fossil members of the Mygalo- morphae in Dominican amber were described by WUNDERLICH (1988). -

(Araneae: Araneomorphae) Antrópicas De Morelia, Michoacán, México
ACAROLOGÍA Y ARACNOLOGÍA ISSN: 2448-475X ARAÑAS (ARANEAE: ARANEOMORPHAE) ANTRÓPICAS DE MORELIA, MICHOACÁN, MÉXICO Juan Maldonado-Carrizales, Ana Quijano-Ravell, Carlos Ernesto Guzmán-García y Javier Ponce-Saavedra 1Laboratorio de Entomología “Biol. Sócrates Cisneros Paz”. Facultad de Biología. UMSNH. Edificio B4 2º. Piso. Ciudad Universitaria, Morelia, Michoacán. Autor de correspondencia: [email protected] RESUMEN. El trabajo se desarrolló en la ciudad de Morelia, Michoacán, México, en 12 puntos de muestreo ubicados en cuatro direcciones partiendo del centro de la ciudad, con tres puntos en cada dirección, orientados en los cuatro puntos cardinales. Se hicieron colectas diurnas en septiembre de 2016 y abril 2017, utilizando colecta directa en el interior y peridomicilio de las casas y 2 horas/persona como unidad de esfuerzo. Se recolectaron 635 organismos, pertenecientes a 20 familias, 29 géneros, seis especies y 29 morfoespecies. El 59.6 % de los ejemplares se recolectaron en el interior de las casas; sin embargo, el índice de diversidad no tuvo diferencia estadística significativa con el peridomicilio, pero sí hubo diferencia en la composición de especies y géneros dominantes. Filistatinella domestica Desales-Lara (Filistatidae) fue la más abundante en ambos sitios y también representa un nuevo registro para el estado de Michoacán. Las estimaciones de riqueza esperada muestran que se requiere de mayor esfuerzo de muestreo, aunque se tuvo una buena representación de la riqueza específica con alrededor del 65 % del número estimado por el modelo de Chao2 Palabras clave: Ecología, sinantropismo, diversidad Anthropic spiders (Araneae: Araneomorphae) of Morelia, Michoacán, México. ABSTRACT. The work was made in Morelia, Michoacan, Mexico, city with 12 sampling points positioned into four directions starting from the downtown. -

Song Dissertation
SYSTEMATICS OF CYRTACANTHACRIDINAE (ORTHOPTERA: ACRIDIDAE) WITH A FOCUS ON THE GENUS SCHISTOCERCA STÅL 1873: EVOLUTION OF LOCUST PHASE POLYPHENISM AND STUDY OF INSECT GENITALIA DISSERTATION Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Hojun Song, M.S. ***** The Ohio State University 2006 Dissertation Committee: Approved by Dr. John W. Wenzel, Advisor Dr. Norman F. Johnson ______________________________ Dr. Johannes S. H. Klompen Advisor Graduate Program in Entomology Copyright by Hojun Song 2006 ABSTRACT The systematics of Cyrtacanthacridinae (Orthoptera: Acrididae) is investigated to study the evolution of locust phase polyphenism, biogeography, and the evolution of male genitalia. In Chapter Two, I present a comprehensive taxonomic synopsis of the genus Schistocerca Stål. I review the taxonomic history, include an identification key to species, revise the species concepts of six species and describe a new species. In Chapter Three, I present a morphological phylogeny of Schistocerca, focusing on the biogeography. The phylogeny places the desert locust S. gregaria deep within the New World clade, suggesting that the desert locust originated from the New World. In Chapter Four, I review the systematics of Cyrtacanthacridinae and present a phylogeny based on morphology. Evolution of taxonomically important characters is investigated using a character optimization analysis. The biogeography of the subfamily is also addressed. In Chapter Five, I present a comprehensive review the recent advances in the study of locust phase polyphenism from various disciplines. The review reveals that locust phase polyphenism is a complex phenomenon consisting of numerous density-dependent phenotypically plastic traits. -

Section III - Acknowledgements
Section III - Acknowledgements Mike Berger TPWD Wildlife Division Director Larry McKinney TPWD Coastal Fisheries Director Phil Durocher TPWD Inland Fisheries Director Lydia Saldana TPWD Communications Division Director TPWD Program Director, Science Research and Diversity Program - Ron George Wildlife Division (Retired) Technical Assistance Andy Price Texas Parks and Wildlife - Wildlife Division Bob Gottfried Texas Parks and Wildlife - Wildlife Division Cliff Shackelford Texas Parks and Wildlife - Wildlife Division Duane Schlitter Texas Parks and Wildlife - Wildlife Division Gary Garrett Texas Parks and Wildlife - Inland Fisheries Division John Young Texas Parks and Wildlife - Wildlife Division Mike Quinn Texas Parks and Wildlife - Wildlife Division Paul Hammerschmidt Texas Parks and Wildlife - Coastal Fisheries Division (Retired) Wildlife Diversity Policy Advisory Committee Terry Austin (chair) Audubon Texas (retired) Damon Waitt Brown Center for Environmental Education - Lady Bird Johnson Wildflower Center David Wolfe Environmental Defense Don Petty Texas Farm Bureau Doug Slack Texas A & M University, Department of Wildlife & Fisheries Sciences Evelyn Merz Lone Star Chapter of the Sierra Club Jack King Sportsmen’s Conservationists of Texas Jennifer Walker Lone Star Chapter of the Sierra Club Jim Bergan The Nature Conservancy of Texas Jim Foster Trans-Texas, Davis Mountain and Hill Country Heritage Association Kirby Brown Texas Wildlife Association Matt Brockman Texas and Southwestern Cattleraisers Association Mike McMurry Texas Department of Agriculture Phil Sudman Texas Society of Mammalogists Richard Egg Texas Soil and Water Conservation Board Susan Kaderka National Wildlife Federation, Gulf States Field Office Ted Eubanks Fermata, Inc. Troy Hibbitts Texas Herpetological Society Wallace Rogers TPWD Private Lands Advisory Board Comprehensive Wildlife Conservation Strategy Working Groups Aquatic Working Group Gary Garrett (chair) Texas Parks and Wildlife Department Paul Hammerschmidt (chair) Texas Parks and Wildlife Department 578 Carrie Thompson U.S. -

Ennominae 70.205 1884 Magpie Moth (Abraxas Grossulariata
Ennominae 70.205 1884 Magpie Moth (Abraxas grossulariata) 70.206 1885 Clouded Magpie (Abraxas sylvata) 70.207 1887 Clouded Border (Lomaspilis marginata) 70.208 1888 Scorched Carpet (Ligdia adustata) 70.210 1888a Dorset Cream Wave (Stegania trimaculata) 70.211 1889 Peacock Moth (Macaria notata) 70.212 1890 Sharp-angled Peacock (Macaria alternata) 70.213 1891 Dusky Peacock (Macaria signaria) 70.214 1893 Tawny-barred Angle (Macaria liturata) 70.215 1897 V-Moth (Macaria wauaria) 70.217 1896 Rannoch Looper (Macaria brunneata) 70.218 1894 Latticed Heath (Chiasmia clathrata) 70.220 1899 Frosted Yellow (Isturgia limbaria) 70.222 1902 Brown Silver-line (Petrophora chlorosata) 70.223 1903 Barred Umber (Plagodis pulveraria) 70.224 1904 Scorched Wing (Plagodis dolabraria) 70.225 1905 Horse Chestnut (Pachycnemia hippocastanaria) 70.226 1906 Brimstone Moth (Opisthograptis luteolata) 70.227 1907 Bordered Beauty (Epione repandaria) 70.228 1908 Dark Bordered Beauty (Epione vespertaria) 70.229 1909 Speckled Yellow (Pseudopanthera macularia) 70.230 1924 Orange Moth (Angerona prunaria) 70.231 1910 Lilac Beauty (Apeira syringaria) 70.232 1911 Large Thorn (Ennomos autumnaria) 70.233 1912 August Thorn (Ennomos quercinaria) 70.234 1913 Canary-shouldered Thorn (Ennomos alniaria) 70.235 1914 Dusky Thorn (Ennomos fuscantaria) 70.236 1915 September Thorn (Ennomos erosaria) 70.237 1917 Early Thorn (Selenia dentaria) 70.238 1918 Lunar Thorn (Selenia lunularia) 70.239 1919 Purple Thorn (Selenia tetralunaria) 70.240 1920 Scalloped Hazel (Odontopera bidentata) 70.241