Genomic Analyses of PMBL Reveal New Drivers and Mechanisms of Sensitivity to PD-1 Blockade
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CIITA Stimulation of Transcription Factor Binding to Major Histocompatibility Complex Class II and Associated Promoters in Vivo
Proc. Natl. Acad. Sci. USA Vol. 95, pp. 6267–6272, May 1998 Genetics CIITA stimulation of transcription factor binding to major histocompatibility complex class II and associated promoters in vivo i KENNETH L. WRIGHT*†§,KEH-CHUANG CHIN‡§¶,MICHAEL LINHOFF*, CHERYL SKINNER*, JEFFREY A. BROWN , i JEREMY M. BOSS ,GEORGE R. STARK**, AND JENNY P.-Y. TING*†† *Lineberger Comprehensive Cancer Center and the Department of Immunology and Microbiology, and ‡Department of Biochemistry and Biophysics, University of North Carolina, Chapel Hill, NC 27599; iDepartment of Microbiology and Immunology, Emory University School of Medicine, Atlanta, GA 30322; and **Lerner Research Institute, The Cleveland Clinic Foundation, 9500 Euclid Avenue, Cleveland, OH 44195 Contributed by George R. Stark, March 12, 1998 ABSTRACT CIITA is a master transactivator of the ma- RFX5 (14). However, the mode of action of CIITA and its in jor histocompatibility complex class II genes, which are vivo target have remained elusive. involved in antigen presentation. Defects in CIITA result in Changes in promoter assembly or proteinyDNA interactions fatal immunodeficiencies. CIITA activation is also the control can be detected in intact cells by in vivo genomic footprinting point for the induction of major histocompatibility complex (15). Analysis of the DRA promoter by this technique has class II and associated genes by interferon-g, but CIITA does revealed interactions at promoter-proximal transcription fac- not bind directly to DNA. Expression of CIITA in G3A cells, tor binding sites X and Y in both B lymphocytes and IFN-g- which lack endogenous CIITA, followed by in vivo genomic treated cells (16, 17). The Ii and DMB promoters show similar footprinting, now reveals that CIITA is required for the interactions at the their X and Y sites (18–20). -

Significant Shortest Paths for the Detection of Putative Disease Modules
bioRxiv preprint doi: https://doi.org/10.1101/2020.04.01.019844; this version posted April 2, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. SIGNIFICANT SHORTEST PATHS FOR THE DETECTION OF PUTATIVE DISEASE MODULES Daniele Pepe1 1Department of Oncology, KU Leuven, LKI–Leuven Cancer Institute, Leuven, Belgium Email address: DP: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/2020.04.01.019844; this version posted April 2, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Keywords Structural equation modeling, significant shortest paths, pathway analysis, disease modules. Abstract Background The characterization of diseases in terms of perturbated gene modules was recently introduced for the analysis of gene expression data. Some approaches were proposed in literature, but many times they are inductive approaches. This means that starting directly from data, they try to infer key gene networks potentially associated to the biological phenomenon studied. However they ignore the biological information already available to characterize the gene modules. Here we propose the detection of perturbed gene modules using the combination of data driven and hypothesis-driven approaches relying on biological metabolic pathways and significant shortest paths tested by structural equation modeling. -

ATP-Binding and Hydrolysis in Inflammasome Activation
molecules Review ATP-Binding and Hydrolysis in Inflammasome Activation Christina F. Sandall, Bjoern K. Ziehr and Justin A. MacDonald * Department of Biochemistry & Molecular Biology, Cumming School of Medicine, University of Calgary, 3280 Hospital Drive NW, Calgary, AB T2N 4Z6, Canada; [email protected] (C.F.S.); [email protected] (B.K.Z.) * Correspondence: [email protected]; Tel.: +1-403-210-8433 Academic Editor: Massimo Bertinaria Received: 15 September 2020; Accepted: 3 October 2020; Published: 7 October 2020 Abstract: The prototypical model for NOD-like receptor (NLR) inflammasome assembly includes nucleotide-dependent activation of the NLR downstream of pathogen- or danger-associated molecular pattern (PAMP or DAMP) recognition, followed by nucleation of hetero-oligomeric platforms that lie upstream of inflammatory responses associated with innate immunity. As members of the STAND ATPases, the NLRs are generally thought to share a similar model of ATP-dependent activation and effect. However, recent observations have challenged this paradigm to reveal novel and complex biochemical processes to discern NLRs from other STAND proteins. In this review, we highlight past findings that identify the regulatory importance of conserved ATP-binding and hydrolysis motifs within the nucleotide-binding NACHT domain of NLRs and explore recent breakthroughs that generate connections between NLR protein structure and function. Indeed, newly deposited NLR structures for NLRC4 and NLRP3 have provided unique perspectives on the ATP-dependency of inflammasome activation. Novel molecular dynamic simulations of NLRP3 examined the active site of ADP- and ATP-bound models. The findings support distinctions in nucleotide-binding domain topology with occupancy of ATP or ADP that are in turn disseminated on to the global protein structure. -

A Role for the Nlr Family Members Nlrc4 and Nlrp3 in Astrocytic Inflammasome Activation and Astrogliosis
A ROLE FOR THE NLR FAMILY MEMBERS NLRC4 AND NLRP3 IN ASTROCYTIC INFLAMMASOME ACTIVATION AND ASTROGLIOSIS Leslie C. Freeman A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Curriculum of Genetics and Molecular Biology. Chapel Hill 2016 Approved by: Jenny P. Y. Ting Glenn K. Matsushima Beverly H. Koller Silva S. Markovic-Plese Pauline. Kay Lund ©2016 Leslie C. Freeman ALL RIGHTS RESERVED ii ABSTRACT Leslie C. Freeman: A Role for the NLR Family Members NLRC4 and NLRP3 in Astrocytic Inflammasome Activation and Astrogliosis (Under the direction of Jenny P.Y. Ting) The inflammasome is implicated in many inflammatory diseases but has been primarily studied in the macrophage-myeloid lineage. Here we demonstrate a physiologic role for nucleotide-binding domain, leucine-rich repeat, CARD domain containing 4 (NLRC4) in brain astrocytes. NLRC4 has been primarily studied in the context of gram-negative bacteria, where it is required for the maturation of pro-caspase-1 to active caspase-1. We show the heightened expression of NLRC4 protein in astrocytes in a cuprizone model of neuroinflammation and demyelination as well as human multiple sclerotic brains. Similar to macrophages, NLRC4 in astrocytes is required for inflammasome activation by its known agonist, flagellin. However, NLRC4 in astrocytes also mediate inflammasome activation in response to lysophosphatidylcholine (LPC), an inflammatory molecule associated with neurologic disorders. In addition to NLRC4, astrocytic NLRP3 is required for inflammasome activation by LPC. Two biochemical assays show the interaction of NLRC4 with NLRP3, suggesting the possibility of a NLRC4-NLRP3 co-inflammasome. -

The RFX Complex Is Crucial for the Constitutive and CIITA-Mediated Transactivation of MHC Class I and 2-Microglobulin Genes
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Immunity, Vol. 9, 531±541, October, 1998, Copyright 1998 by Cell Press The RFX Complex Is Crucial for the Constitutive and CIITA-Mediated Transactivation of MHC Class I and b2-Microglobulin Genes Sam J. P. Gobin,² Ad Peijnenburg,² of the NF-kB/Rel family and are thought to be essential Marja van Eggermond, Marlijn van Zutphen, for both constitutive and cytokine-induced expression Rian van den Berg, and Peter J. van den Elsen* (Girdlestone et al., 1993; Mansky et al., 1994). The ISRE Department of Immunohematology and Blood Bank is bound by factors of the interferon regulatory factor Leiden University Medical Center (IRF) family and mediates the induction of MHC class I 2333 ZA Leiden expression by interferons (Girdlestone et al., 1993). Site The Netherlands a was originally described as being important for the constitutive expression of MHC class I genes (Dey et al., 1992). However, a crucial role in a novel route of Summary HLA class I gene activation mediated by the class II transactivator (CIITA) has recently been attributed to In type III bare lymphocyte syndrome (BLS) patients, site a (Gobin et al., 1997; Martin et al., 1997). Enhancer defects in the RFX protein complex result in a lack of B contains an inverted CCAAT box sequence that plays MHC class II and reduced MHC class I cell surface a role in basal MHC class I gene transcription (Schoneich et al., 1997). In the promoter region of the m gene, expression. -

Transcriptome Analysis of Human Diabetic Kidney Disease
ORIGINAL ARTICLE Transcriptome Analysis of Human Diabetic Kidney Disease Karolina I. Woroniecka,1 Ae Seo Deok Park,1 Davoud Mohtat,2 David B. Thomas,3 James M. Pullman,4 and Katalin Susztak1,5 OBJECTIVE—Diabetic kidney disease (DKD) is the single cases, mild and then moderate mesangial expansion can be leading cause of kidney failure in the U.S., for which a cure has observed. In general, diabetic kidney disease (DKD) is not yet been found. The aim of our study was to provide an considered a nonimmune-mediated degenerative disease unbiased catalog of gene-expression changes in human diabetic of the glomerulus; however, it has long been noted that kidney biopsy samples. complement and immunoglobulins sometimes can be de- — tected in diseased glomeruli, although their role and sig- RESEARCH DESIGN AND METHODS Affymetrix expression fi arrays were used to identify differentially regulated transcripts in ni cance is not clear (4). 44 microdissected human kidney samples. The DKD samples were The understanding of DKD has been challenged by multi- significant for their racial diversity and decreased glomerular ple issues. First, the diagnosis of DKD usually is made using filtration rate (~20–30 mL/min). Stringent statistical analysis, using clinical criteria, and kidney biopsy often is not performed. the Benjamini-Hochberg corrected two-tailed t test, was used to According to current clinical practice, the development of identify differentially expressed transcripts in control and diseased albuminuria in patients with diabetes is sufficient to make the glomeruli and tubuli. Two different Web-based algorithms were fi diagnosis of DKD (5). We do not understand the correlation used to de ne differentially regulated pathways. -

DCIR2 Cdc2 Dcs and Zbtb32 Restore CD4 T-Cell Tolerance
Diabetes Volume 64, October 2015 3521 Jeffrey D. Price, Chie Hotta-Iwamura, Yongge Zhao, Nicole M. Beauchamp, and Kristin V. Tarbell DCIR2+ cDC2 DCs and Zbtb32 Restore CD4+ T-Cell Tolerance and Inhibit Diabetes Diabetes 2015;64:3521–3531 | DOI: 10.2337/db14-1880 During autoimmunity, the normal ability of dendritic cells clinical efficacy has been observed for this approach (1,2). (DCs) to induce T-cell tolerance is disrupted; therefore, Autoimmune individuals elicit immune responses in an autoimmune disease therapies based on cell types and inflammatory context and are therefore refractory to tol- IMMUNOLOGY AND TRANSPLANTATION molecular pathways that elicit tolerance in the steady erance induction, yet most studies of T-cell tolerance have state may not be effective. To determine which DC been performed in either a steady-state context or in subsets induce tolerance in the context of chronic models of autoimmunity requiring immunization with fi autoimmunity, we used chimeric antibodies speci cfor autoantigen that best model the effector phase (3). There- DC inhibitory receptor 2 (DCIR2) or DEC-205 to target fi + + fore, to move beyond therapies that nonspeci cally block self-antigen to CD11b (cDC2) DCs and CD8 (cDC1) DCs, effector functions, it is important to learn what condi- respectively, in autoimmune-prone nonobese diabetic tions are needed to enable antigen-specific T-cell tolerance (NOD) mice. Antigen presentation by DCIR2+ DCs but induction in a chronic inflammatory autoimmune envi- not DEC-205+ DCs elicited tolerogenic CD4+ T-cell ronment, which can be modeled using autoimmune-prone responses in NOD mice. b-Cell antigen delivered to DCIR2+ DCs delayed diabetes induction and induced in- nonobese diabetic (NOD) mice that show spontaneous creased T-cell apoptosis without interferon-g (IFN-g)or loss of self-tolerance due to genetic and environmental sustained expansion of autoreactive CD4+ T cells. -
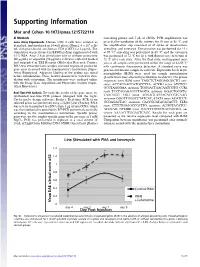
Supporting Information
Supporting Information Mor and Cohen 10.1073/pnas.1215722110 SI Methods containing primer and 5 μL of cDNA. PCR amplification was Gene Array Experiments. Human CD4 T cells were isolated as preceded by incubation of the mixture for 10 min at 95 °C, and described, and incubated in 24-well plates (Nunc), 4 × 106 cells/ the amplification step consisted of 45 cycles of denaturation, mL with plate-bound anti-human CD3 (OKT3) at 2 μg/mL. The annealing, and extension. Denaturation was performed for 15 s stimulation was performed in RPMI medium supplemented with at 95 °C; annealing was performed in 60 °C; and the extension 0.1% BSA. After 2 h of stimulation with or without cefuroxime was performed at 72 °C for 20 s, with fluorescence detection at (50 μg/mL) or ampicillin (50 μg/mL), cells were collected washed 72 °C after each cycle. After the final cycle, melting-point anal- and suspended in TRI Reagent (Molecular Research Center). yses of all samples were performed within the range of 62–95 °C RNA was extracted from samples and used to prepare probes for with continuous fluorescence detection. A standard curve was gene array in accord with the manufacturer’s instructions (Super- generated from one sample in each run. Expression levels of β2- Array Bioscience). Adequate labeling of the probes was tested microglobulin (B2M) were used for sample normalization before hybridization. Three healthy donors were tested in stim- (β-actin levels were affected by cefuroxime treatment). The primer ulation with cefuroxime. The membranes were analyzed online sequences were B2M sense TAGCTCTAGGAGGGCTG anti- with the Image Data Acquisition and Expression Analysis (Super- sense ACCACAACCATGCCTTA; ACVR2 sense ATCTCC- Array Bioscience). -

Genome-Wide Identification of Genes Regulating DNA Methylation Using Genetic Anchors for Causal Inference
bioRxiv preprint doi: https://doi.org/10.1101/823807; this version posted October 30, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. Genome-wide identification of genes regulating DNA methylation using genetic anchors for causal inference Paul J. Hop1,2, René Luijk1, Lucia Daxinger3, Maarten van Iterson1, Koen F. Dekkers1, Rick Jansen4, BIOS Consortium5, Joyce B.J. van Meurs6, Peter A.C. ’t Hoen 7, M. Arfan Ikram8, Marleen M.J. van Greevenbroek9,10, Dorret I. Boomsma11, P. Eline Slagboom1, Jan H. Veldink2, Erik W. van Zwet12, Bastiaan T. Heijmans1* 1 Molecular Epidemiology, Department of Biomedical Data Sciences, Leiden University Medical Center, Leiden, 2333 ZC, The Netherlands 2 Department of Neurology, UMC Utrecht Brain Center, University Medical Centre Utrecht, Utrecht University, Utrecht 3584 CG, Netherlands 3 Department of Human Genetics, Leiden University Medical Center, Leiden, 2333 ZC, The Netherlands 4 Department of Psychiatry, Amsterdam UMC, Vrije Universiteit Amsterdam, Amsterdam Neuroscience, Amsterdam, 1081 HV, The Netherlands 5 Biobank-based Integrated Omics Study Consortium. For a complete list of authors, see the acknowledgements. 6 Department of Internal Medicine, Erasmus Medical Centre, Rotterdam, The Netherlands. 7 Centre for Molecular and Biomolecular Informatics, Radboud Institute for Molecular Life Sciences, Radboud University -

The Role of the NFAT Signalling Pathway in Diffuse Large B-Cell Lymphoma
The role of the NFAT signalling pathway in Diffuse Large B-cell Lymphoma Holly White Doctor of Philosophy Thesis September 2015 Supervisors: Dr Chris Bacon, Professor Neil Perkins & Dr Vikki Rand Northern Institute for Cancer Research Word count: 66,024 1 Abstract Diffuse Large B-Cell Lymphomas (DLBCL) are common, aggressive malignancies of mature B-lymphocytes that represent ~40% of lymphomas. Despite the widespread use of combined immunochemotherapy, approximately 50% of patients with DLBCL die from their disease. The two main DLBCL subgroups resemble activated B cells (ABC) or germinal centre B cells (GCB), where patients with ABC-DLBCL have significantly worse outcome. There is urgent need for novel therapeutic strategies in the treatment of DLBCL, which requires a better understanding of the molecular pathways upon which tumours depend. Accumulating evidence suggests that the signalling networks promoting and sustaining DLBCL derive from dysregulation of the normal pathways controlling B-lymphocyte activation and differentiation. There is increasing evidence indicating important roles for the NFAT family of transcription factors in DLBCL. Constitutively-active nuclear NFAT2 has been demonstrated in approximately 40% of primary DLBCL samples and NFAT has been shown to regulate a small number of genes associated with DLBCL growth/survival. This project investigated the role of NFAT in DLBCL. Nuclear localisation and activation of NFAT family members were characterised in a panel of DLBCL cell lines and chemical inhibition of calcineurin/NFAT, using Cyclosporin A (CsA), indicated dependency on the calcineurin/NFAT pathway for survival. Gene expression microarray analysis performed in DLBCL cell lines treated with CsA revealed potential NFAT target genes involved in the tumour microenvironment and anergy. -

Recurrent Genetic Mutations in Lymphoid Malignancies
Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1312 Recurrent Genetic Mutations in Lymphoid Malignancies EMMA YOUNG ACTA UNIVERSITATIS UPSALIENSIS ISSN 1651-6206 ISBN 978-91-554-9850-4 UPPSALA urn:nbn:se:uu:diva-314956 2017 Dissertation presented at Uppsala University to be publicly examined in Rudbecksalen, Dag Hammarskjölds Väg 20, Uppsala, Friday, 5 May 2017 at 13:15 for the degree of Doctor of Philosophy (Faculty of Medicine). The examination will be conducted in English. Faculty examiner: Professor Dimitar Efremov (Molecular Hematology Group of the International Centre for Genetic Engineering and Biotechnology (ICGEB) in Trieste, Italy). Abstract Young, E. 2017. Recurrent Genetic Mutations in Lymphoid Malignancies. Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1312. 82 pp. Uppsala: Acta Universitatis Upsaliensis. ISBN 978-91-554-9850-4. In recent years, the genetic landscape of B-cell derived lymphoid malignancies, including chronic lymphocytic leukemia (CLL), has been rapidly unraveled, identifying recurrent genetic mutations with potential clinical impact. Interestingly, ~30% of all CLL patients can be assigned to more homogeneous subsets based on the expression of a similar or “stereotyped” B-cell receptor (BcR). Considering that biased distribution of genetic mutations was recently indicated in specific stereotyped subsets, in paper I, we screened 565 subset cases, preferentially assigned to clinically aggressive subsets, and confirm the SF3B1 mutational bias in subset #2 (45%), but also report on similarly marked enrichment in subset #3 (46%). In contrast, NOTCH1 mutations were predominantly detected in subsets #1, #8, #59 and #99 (22-34%). This data further highlights a subset-biased acquisition of genetic mutations in the pathogenesis of at least certain subsets. -

Supplemental Materials
SUPPLEMENTARY DATA shRNA-containing lentiviral vectors. The shRNAs for each target genes were designed with the aid of web-based Invitrogen Block-It program. The oligonucleotides containing Fra-1, c-Jun or slug shRNA sequences were inserted into pLV-shRNA vector (Biosettia, San Diego, CA) according to manufacturer’s protocol. The Fra-1 mRNA target sequences are 5’- GGAGACTGACAAACTGGAAGA-3’ and 5’-GGATGGTACAGCCTCATTTCC-3’. The c-Jun mRNA target sequences are 5’-GGAACAGGTGGCACAGCTTAA-3’ and 5’- GGCACAGCTTAAACAGAAAGT-3’. The slug mRNA target sequences are 5’- GCATTTGCAGACAGGTCAAAT-3’ and 5’-GCTCATCTGCAGACCCATTCT-3’. Slug promoter construction. The slug promoter (GeneBank Accession number AB300659) was synthesized by PCR using genomic DNA isolated from MDA-MB-231 cells. The synthesized fragment was subcloned into pGL4-luciferase reporter gene plasmid (Promega, Madison, WI) and sequence confirmed by automated DNA sequencing. To generate AP1 mutant, the AP1 consensus sequence in the slug promoter was destroyed by introducing mutation at nucleotide 1009-1011 (GAC AGA). To determine the promoter activity, wild-type or mutant slug promoter reporter gene construct was transfected into MCF-7, MDA-MB-231, MDA-MB-436 or ZR-75-1 cells for 36 hrs and cell lysates collected to measure luciferase activity using Lipofectamine (Invitrogen, Carlsbad, CA). To determine the effect of Fra-1/c-Jun expression on slug promoter activity, the promoter reporter gene construct was co-transfected with expression vectors encoding Fra-1 and c-Jun into MCF-7 and ZR-75-1 cells for 48 hrs followed by the analysis of luciferase activity. Plasmid containing Renilla Luciferase gene was included during transfection to serve as an internal control and Dual Luciferase Assay System (Promega, Madison, WI) was used for measuring luciferase activity.