The Diversity of Intermediate Filaments in Astrocytes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Role of Nuclear Lamin B1 in Cell Proliferation and Senescence
Downloaded from genesdev.cshlp.org on September 29, 2021 - Published by Cold Spring Harbor Laboratory Press The role of nuclear lamin B1 in cell proliferation and senescence Takeshi Shimi,1 Veronika Butin-Israeli,1 Stephen A. Adam,1 Robert B. Hamanaka,2 Anne E. Goldman,1 Catherine A. Lucas,1 Dale K. Shumaker,1 Steven T. Kosak,1 Navdeep S. Chandel,2 and Robert D. Goldman1,3 1Department of Cell and Molecular Biology, 2Department of Medicine, Division of Pulmonary and Critical Care Medicine, Feinberg School of Medicine, Northwestern University, Chicago, Illinois 60611, USA Nuclear lamin B1 (LB1) is a major structural component of the nucleus that appears to be involved in the regulation of many nuclear functions. The results of this study demonstrate that LB1 expression in WI-38 cells decreases during cellular senescence. Premature senescence induced by oncogenic Ras also decreases LB1 expression through a retinoblastoma protein (pRb)-dependent mechanism. Silencing the expression of LB1 slows cell proliferation and induces premature senescence in WI-38 cells. The effects of LB1 silencing on proliferation require the activation of p53, but not pRb. However, the induction of premature senescence requires both p53 and pRb. The proliferation defects induced by silencing LB1 are accompanied by a p53-dependent reduction in mitochondrial reactive oxygen species (ROS), which can be rescued by growth under hypoxic conditions. In contrast to the effects of LB1 silencing, overexpression of LB1 increases the proliferation rate and delays the onset of senescence of WI-38 cells. This overexpression eventually leads to cell cycle arrest at the G1/S boundary. -
The Desmoplakin Carboxyl Terminus Coaligns with and Specifically Disrupts Intermediate Filament Networks When Expressed in Cultured Cells Thaddeus S
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by PubMed Central The Desmoplakin Carboxyl Terminus Coaligns with and Specifically Disrupts Intermediate Filament Networks When Expressed in Cultured Cells Thaddeus S. Stappenbeck and Kathleen J. Green Department of Pathology and the Cancer Center, Northwestern University Medical School, Chicago, Illinois 60611 Abstract. Specific interactions between desmoplakins tides including the 90-kD carboxy-terminal globular I and 11 (DP I and II) and other desmosomal or cyto- domain of DP I specifically colocalized with and ulti- skeletal molecules have been difficult to determine in mately resulted in the complete disruption of IF in part because of the complexity and insolubility of the both cell lines. This effect was specific for IF as micro- desmosome and its constituents . We have used a mo- tubule and microfilament networks were unaltered . lecular genetic approach to investigate the role that This effect was also specific for the carboxyl terminus DP I and 11 may play in the association of the desmo- of DP, as the expression of the 95-kD rod domain of somal plaque with cytoplasmic intermediate filaments DP I did not visibly alter IF networks. Immunogold (IF) . A series of mammalian expression vectors en- localization of COS-7 cells transfected with constructs coding specific predicted domains of DP I were tran- including the carboxyl terminus of DP demonstrated siently expressed in cultured cells that form (COS-7) an accumulation of mutant protein in perinuclear aggre- and do not form (NIH-3T3) desmosomes. Sequence gates within which IF subunits were sequestered. -

Blood Neurofilament Light Chain: the Neurologist's Troponin?
biomedicines Review Blood Neurofilament Light Chain: The Neurologist’s Troponin? Simon Thebault 1,*, Ronald A. Booth 2 and Mark S. Freedman 1,* 1 Department of Medicine and the Ottawa Hospital Research Institute, The University of Ottawa, Ottawa, ON K1H8L6, Canada 2 Department of Pathology and Laboratory Medicine, Eastern Ontario Regional Laboratory Association and Ottawa Hospital Research Institute, University of Ottawa & The Ottawa Hospital, Ottawa, ON K1H8L6, Canada; [email protected] * Correspondence: [email protected] (S.T.); [email protected] (M.S.F.) Received: 4 November 2020; Accepted: 18 November 2020; Published: 21 November 2020 Abstract: Blood neurofilament light chain (NfL) is a marker of neuro-axonal injury showing promising associations with outcomes of interest in several neurological conditions. Although initially discovered and investigated in the cerebrospinal fluid (CSF), the recent development of ultrasensitive digital immunoassay technologies has enabled reliable detection in serum/plasma, obviating the need for invasive lumbar punctures for longitudinal assessment. The most evidence for utility relates to multiple sclerosis (MS) where it serves as an objective measure of both the inflammatory and degenerative pathologies that characterise this disease. In this review, we summarise the physiology and pathophysiology of neurofilaments before focusing on the technological advancements that have enabled reliable quantification of NfL in blood. As the test case for clinical translation, we then highlight important recent developments linking blood NfL levels to outcomes in MS and the next steps to be overcome before this test is adopted on a routine clinical basis. Keywords: neurofilament light chain; biomarkers; multiple sclerosis 1. Neurofilament Structure and Function Neurofilaments are neuronal-specific heteropolymers conventionally considered to consist of a triplet of light (NfL), medium (NfM) and heavy (NfH) chains according to their molecular mass [1]. -

Haploinsufficiency of the Schizophrenia and Autism Risk Gene
Haan et al. Translational Psychiatry (2021) 11:313 https://doi.org/10.1038/s41398-021-01415-6 Translational Psychiatry ARTICLE Open Access Haploinsufficiency of the schizophrenia and autism risk gene Cyfip1 causes abnormal postnatal hippocampal neurogenesis through microglial and Arp2/3 mediated actin dependent mechanisms Niels Haan 1,LauraJ.Westacott1, Jenny Carter1,MichaelJ.Owen 1, William P. Gray1,2,3, Jeremy Hall 1,3 and Lawrence S. Wilkinson1,3,4 Abstract Genetic risk factors can significantly increase chances of developing psychiatric disorders, but the underlying biological processes through which this risk is effected remain largely unknown. Here we show that haploinsufficiency of Cyfip1, a candidate risk gene present in the pathogenic 15q11.2(BP1–BP2) deletion may impact on psychopathology via abnormalities in cell survival and migration of newborn neurons during postnatal hippocampal neurogenesis. We demonstrate that haploinsufficiency of Cyfip1 leads to increased numbers of adult-born hippocampal neurons due to reduced apoptosis, without altering proliferation. We show this is due to a cell autonomous failure of microglia to induce apoptosis through the secretion of the appropriate factors, a previously undescribed mechanism. Furthermore, we show an abnormal migration of adult-born neurons due to altered Arp2/3 mediated actin dynamics. Together, our findings throw new light on how the genetic risk candidate Cyfip1 may influence the hippocampus, a brain region 1234567890():,; 1234567890():,; 1234567890():,; 1234567890():,; with strong evidence for involvement in psychopathology. Introduction (BP1–BP2) psychiatric phenotype due to evidence of Many psychiatric conditions show high heritability. CYFIP1’s involvement in a range of synaptic functions, Recent genetic studies in schizophrenia for example have including key roles in dendritic spine morphology and – identified up to 160 loci that increase risk for the dis- branching9 11. -

The National Economic Burden of Rare Disease Study February 2021
Acknowledgements This study was sponsored by the EveryLife Foundation for Rare Diseases and made possible through the collaborative efforts of the national rare disease community and key stakeholders. The EveryLife Foundation thanks all those who shared their expertise and insights to provide invaluable input to the study including: the Lewin Group, the EveryLife Community Congress membership, the Technical Advisory Group for this study, leadership from the National Center for Advancing Translational Sciences (NCATS) at the National Institutes of Health (NIH), the Undiagnosed Diseases Network (UDN), the Little Hercules Foundation, the Rare Disease Legislative Advocates (RDLA) Advisory Committee, SmithSolve, and our study funders. Most especially, we thank the members of our rare disease patient and caregiver community who participated in this effort and have helped to transform their lived experience into quantifiable data. LEWIN GROUP PROJECT STAFF Grace Yang, MPA, MA, Vice President Inna Cintina, PhD, Senior Consultant Matt Zhou, BS, Research Consultant Daniel Emont, MPH, Research Consultant Janice Lin, BS, Consultant Samuel Kallman, BA, BS, Research Consultant EVERYLIFE FOUNDATION PROJECT STAFF Annie Kennedy, BS, Chief of Policy and Advocacy Julia Jenkins, BA, Executive Director Jamie Sullivan, MPH, Director of Policy TECHNICAL ADVISORY GROUP Annie Kennedy, BS, Chief of Policy & Advocacy, EveryLife Foundation for Rare Diseases Anne Pariser, MD, Director, Office of Rare Diseases Research, National Center for Advancing Translational Sciences (NCATS), National Institutes of Health Elisabeth M. Oehrlein, PhD, MS, Senior Director, Research and Programs, National Health Council Christina Hartman, Senior Director of Advocacy, The Assistance Fund Kathleen Stratton, National Academies of Science, Engineering and Medicine (NASEM) Steve Silvestri, Director, Government Affairs, Neurocrine Biosciences Inc. -
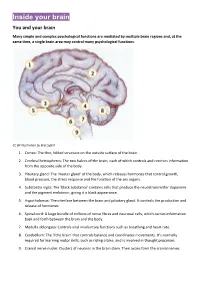
Inside Your Brain You and Your Brain
Inside your brain You and your brain Many simple and complex psychological functions are mediated by multiple brain regions and, at the same time, a single brain area may control many psychological functions. CC BY Illustration by Bret Syfert 1. Cortex: The thin, folded structure on the outside surface of the brain. 2. Cerebral hemispheres: The two halves of the brain, each of which controls and receives information from the opposite side of the body. 3. Pituitary gland: The ‘master gland’ of the body, which releases hormones that control growth, blood pressure, the stress response and the function of the sex organs. 4. Substantia nigra: The ‘black substance’ contains cells that produce the neurotransmitter dopamine and the pigment melatonin, giving it a black appearance. 5. Hypothalamus: The interface between the brain and pituitary gland. It controls the production and release of hormones. 6. Spinal cord: A large bundle of millions of nerve fibres and neuronal cells, which carries information back and forth between the brain and the body. 7. Medulla oblongata: Controls vital involuntary functions such as breathing and heart rate. 8. Cerebellum: The ‘little brain’ that controls balance and coordinates movements. It’s normally required for learning motor skills, such as riding a bike, and is involved in thought processes. 9. Cranial nerve nuclei: Clusters of neurons in the brain stem. Their axons form the cranial nerves. Your brain underpins who you are. It stores your knowledge and memories, gives you the capacity for thought and emotion, and enables you to control your body. The brain is just one part of the nervous system. -

Table 2. Significant
Table 2. Significant (Q < 0.05 and |d | > 0.5) transcripts from the meta-analysis Gene Chr Mb Gene Name Affy ProbeSet cDNA_IDs d HAP/LAP d HAP/LAP d d IS Average d Ztest P values Q-value Symbol ID (study #5) 1 2 STS B2m 2 122 beta-2 microglobulin 1452428_a_at AI848245 1.75334941 4 3.2 4 3.2316485 1.07398E-09 5.69E-08 Man2b1 8 84.4 mannosidase 2, alpha B1 1416340_a_at H4049B01 3.75722111 3.87309653 2.1 1.6 2.84852656 5.32443E-07 1.58E-05 1110032A03Rik 9 50.9 RIKEN cDNA 1110032A03 gene 1417211_a_at H4035E05 4 1.66015788 4 1.7 2.82772795 2.94266E-05 0.000527 NA 9 48.5 --- 1456111_at 3.43701477 1.85785922 4 2 2.8237185 9.97969E-08 3.48E-06 Scn4b 9 45.3 Sodium channel, type IV, beta 1434008_at AI844796 3.79536664 1.63774235 3.3 2.3 2.75319499 1.48057E-08 6.21E-07 polypeptide Gadd45gip1 8 84.1 RIKEN cDNA 2310040G17 gene 1417619_at 4 3.38875643 1.4 2 2.69163229 8.84279E-06 0.0001904 BC056474 15 12.1 Mus musculus cDNA clone 1424117_at H3030A06 3.95752801 2.42838452 1.9 2.2 2.62132809 1.3344E-08 5.66E-07 MGC:67360 IMAGE:6823629, complete cds NA 4 153 guanine nucleotide binding protein, 1454696_at -3.46081884 -4 -1.3 -1.6 -2.6026947 8.58458E-05 0.0012617 beta 1 Gnb1 4 153 guanine nucleotide binding protein, 1417432_a_at H3094D02 -3.13334396 -4 -1.6 -1.7 -2.5946297 1.04542E-05 0.0002202 beta 1 Gadd45gip1 8 84.1 RAD23a homolog (S. -

The Spectraplakin Dystonin Antagonizes YAP Activity and Suppresses Tumourigenesis Praachi B
www.nature.com/scientificreports OPEN The spectraplakin Dystonin antagonizes YAP activity and suppresses tumourigenesis Praachi B. Jain1,2,3, Patrícia S. Guerreiro1,2,3, Sara Canato1,2,3 & Florence Janody 1,2,3* Aberrant expression of the Spectraplakin Dystonin (DST) has been observed in various cancers, including those of the breast. However, little is known about its role in carcinogenesis. In this report, we demonstrate that Dystonin is a candidate tumour suppressor in breast cancer and provide an underlying molecular mechanism. We show that in MCF10A cells, Dystonin is necessary to restrain cell growth, anchorage-independent growth, self-renewal properties and resistance to doxorubicin. Strikingly, while Dystonin maintains focal adhesion integrity, promotes cell spreading and cell-substratum adhesion, it prevents Zyxin accumulation, stabilizes LATS and restricts YAP activation. Moreover, treating DST- depleted MCF10A cells with the YAP inhibitor Verteporfn prevents their growth. In vivo, the Drosophila Dystonin Short stop also restricts tissue growth by limiting Yorkie activity. As the two Dystonin isoforms BPAG1eA and BPAG1e are necessary to inhibit the acquisition of transformed features and are both downregulated in breast tumour samples and in MCF10A cells with conditional induction of the Src proto-oncogene, they could function as the predominant Dystonin tumour suppressor variants in breast epithelial cells. Thus, their loss could deem as promising prognostic biomarkers for breast cancer. Breast cancer progression depends on cell autonomous regulatory mechanisms, driven by mutations and epi- genetic changes, and on non-cell autonomous interactions with the surrounding tumour microenvironment1,2. During this multistep process, normal breast epithelial cells acquire various cellular properties arising from dereg- ulated cellular signalling3,4. -

BD Horizon™ V450 Mouse Anti-Nestin
BD Horizon™ Technical Data Sheet V450 Mouse anti-Nestin Product Information Material Number: 561551 Size: 50 tests Vol. per Test: 5 µl Clone: 25/NESTIN Immunogen: Rat Nestin aa. 402-604 Recombinant Protein Isotype: Mouse IgG1, κ Reactivity: QC Testing: Rat Reported: Human Storage Buffer: Aqueous buffered solution containing protein stabilizer and ≤0.09% sodium azide. Description The cytoskeleton consists primarily of core structural proteins that include microfilaments, microtubules, and intermediate filaments (IFs). IFs contain more than 50 distinct proteins that are organized into six different subtypes: Type I/II keratins expressed in epithelia, type III vimentin/desmin, type IV neurofilament proteins, type V nuclear lamins, and type VI nestin expressed primarily in embryonic cells. Nestin has a conserved core region (amino acids 7 to 314), which contains an α helical domain that is involved in coiled-coil assembly of IFs. The C-terminal region of nestin is similar to type IV IFs, since it contains highly charged amino acids, many glutamate residues, and an 11 amino acid repeat motif. Nestin is expressed in the cerebrum during embryonic development, in the cerebellum during early postnatal development, and in dermatomal cells and myoblasts during myogenesis. In vitro, nestin forms homodimers and homotetramers, but not IFs, and can co-assemble with type III vimentin and type IV internexin proteins. Thus, nestin is a core IF protein that is essential for proper cytoskeletal formation during neurogenesis and myogenesis. The antibody is conjugated to BD Horizon™ V450, which has been developed for use in multicolor flow cytometry experiments and is available exclusively from BD Biosciences. -

Drp1 Overexpression Induces Desmin Disassembling and Drives Kinesin-1 Activation Promoting Mitochondrial Trafficking in Skeletal Muscle
Cell Death & Differentiation (2020) 27:2383–2401 https://doi.org/10.1038/s41418-020-0510-7 ARTICLE Drp1 overexpression induces desmin disassembling and drives kinesin-1 activation promoting mitochondrial trafficking in skeletal muscle 1 1 2 2 2 3 Matteo Giovarelli ● Silvia Zecchini ● Emanuele Martini ● Massimiliano Garrè ● Sara Barozzi ● Michela Ripolone ● 3 1 4 1 5 Laura Napoli ● Marco Coazzoli ● Chiara Vantaggiato ● Paulina Roux-Biejat ● Davide Cervia ● 1 1 2 1,4 6 Claudia Moscheni ● Cristiana Perrotta ● Dario Parazzoli ● Emilio Clementi ● Clara De Palma Received: 1 August 2019 / Revised: 13 December 2019 / Accepted: 23 January 2020 / Published online: 10 February 2020 © The Author(s) 2020. This article is published with open access Abstract Mitochondria change distribution across cells following a variety of pathophysiological stimuli. The mechanisms presiding over this redistribution are yet undefined. In a murine model overexpressing Drp1 specifically in skeletal muscle, we find marked mitochondria repositioning in muscle fibres and we demonstrate that Drp1 is involved in this process. Drp1 binds KLC1 and enhances microtubule-dependent transport of mitochondria. Drp1-KLC1 coupling triggers the displacement of KIF5B from 1234567890();,: 1234567890();,: kinesin-1 complex increasing its binding to microtubule tracks and mitochondrial transport. High levels of Drp1 exacerbate this mechanism leading to the repositioning of mitochondria closer to nuclei. The reduction of Drp1 levels decreases kinesin-1 activation and induces the partial recovery of mitochondrial distribution. Drp1 overexpression is also associated with higher cyclin-dependent kinase-1 (Cdk-1) activation that promotes the persistent phosphorylation of desmin at Ser-31 and its disassembling. Fission inhibition has a positive effect on desmin Ser-31 phosphorylation, regardless of Cdk-1 activation, suggesting that induction of both fission and Cdk-1 are required for desmin collapse. -

Plakoglobin Is Required for Effective Intermediate Filament Anchorage to Desmosomes Devrim Acehan1, Christopher Petzold1, Iwona Gumper2, David D
ORIGINAL ARTICLE Plakoglobin Is Required for Effective Intermediate Filament Anchorage to Desmosomes Devrim Acehan1, Christopher Petzold1, Iwona Gumper2, David D. Sabatini2, Eliane J. Mu¨ller3, Pamela Cowin2,4 and David L. Stokes1,2,5 Desmosomes are adhesive junctions that provide mechanical coupling between cells. Plakoglobin (PG) is a major component of the intracellular plaque that serves to connect transmembrane elements to the cytoskeleton. We have used electron tomography and immunolabeling to investigate the consequences of PG knockout on the molecular architecture of the intracellular plaque in cultured keratinocytes. Although knockout keratinocytes form substantial numbers of desmosome-like junctions and have a relatively normal intercellular distribution of desmosomal cadherins, their cytoplasmic plaques are sparse and anchoring of intermediate filaments is defective. In the knockout, b-catenin appears to substitute for PG in the clustering of cadherins, but is unable to recruit normal levels of plakophilin-1 and desmoplakin to the plaque. By comparing tomograms of wild type and knockout desmosomes, we have assigned particular densities to desmoplakin and described their interaction with intermediate filaments. Desmoplakin molecules are more extended in wild type than knockout desmosomes, as if intermediate filament connections produced tension within the plaque. On the basis of our observations, we propose a particular assembly sequence, beginning with cadherin clustering within the plasma membrane, followed by recruitment of plakophilin and desmoplakin to the plaque, and ending with anchoring of intermediate filaments, which represents the key to adhesive strength. Journal of Investigative Dermatology (2008) 128, 2665–2675; doi:10.1038/jid.2008.141; published online 22 May 2008 INTRODUCTION dense plaque that is further from the membrane and that Desmosomes are large macromolecular complexes that mediates the binding of intermediate filaments. -

Cytoskeletal Linkers: New Maps for Old Destinations Megan K
R864 Dispatch Cytoskeletal linkers: New MAPs for old destinations Megan K. Houseweart*† and Don W. Cleveland*†‡§ A new isoform of the actin–neurofilament linker protein as ‘bullous pemphigoid antigen’ (BPAG). These proteins BPAG has been found that binds to and stabilizes are large α-helical coiled-coil molecules which have axonal microtubules. This and other newly identified binding domains for one or more of the three cytoskele- microtubule-associated proteins are likely to be just the tal components (Figure 1). For example, the widely tip of an iceberg of multifunctional proteins that expressed, > 500 kD protein plectin has been shown to stabilize and crosslink cytoskeletal filament networks. associate with microtubules, intermediate filaments (glial fibrillary acidic protein, vimentin, keratins, all Addresses: *Ludwig Institute for Cancer Research, †Program in Biomedical Sciences, ‡Division of Cellular and Molecular Medicine and three neurofilament subunit proteins), actin, myosin and §Department of Neuroscience, University of California at San Diego, itself [3]. Given the widespread distribution and multi- La Jolla, California 92093, USA. ple interactions that are characteristic of these proteins, E-mail: [email protected] it is not surprising that a number of human and mouse Current Biology 1999, 9:R864–R866 diseases have been attributed to aberrant or missing cross-linking proteins [4]. 0960-9822/99/$ – see front matter © 1999 Elsevier Science Ltd. All rights reserved. This is the case for mice lacking the locus encoding the numerous isoforms of the essential ~280 kDa linker The cytoplasm of most eukaryotic cells contains a dynamic protein BPAG. Two neuronal isoforms of BPAG both have filamentous protein scaffold composed of 25 nm micro- a carboxy-terminal intermediate-filament-binding domain tubules, 4 nm actin filaments and 10 nm intermediate fila- and also an amino-terminal actin-binding region (Figure 1).