Symmetries, Generalized Numbers and Harmonic Laws in Matrix Genetics
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
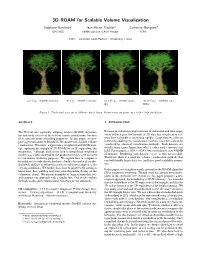
3D ROAM for Scalable Volume Visualization
3D ROAM for Scalable Volume Visualization St´ephane Marchesin∗ Jean-Michel Dischler† Catherine Mongenet‡ ICPS/IGG INRIA Lorraine, CALVI Project ICPS LSIIT { Universit´e Louis Pasteur { Strasbourg, France (a) 2.5 fps – 104000 tetrahedra (b) 1 fps – 380000 tetrahedra (c) 0.45 fps – 932000 tetrahe- (d) 0.15 fps – 3065000 tetra- dra hedra Figure 1: The bonsa¨ı data set at different detail levels. Frame rates are given for a 1024 × 1024 resolution ABSTRACT 1 INTRODUCTION The 2D real time optimally adapting meshes (ROAM) algorithm Because of continuous improvements in simulation and data acqui- has had wide success in the field of terrain visualization, because sition technologies, the amount of 3D data that visualization sys- of its efficient error-controlling properties. In this paper, we pro- tems have to handle is increasing rapidly. Large datasets offer an pose a generalization of ROAM in 3D suitable for scalable volume interesting challenge to visualization systems, since they cannot be visualization. Therefore, we perform a straightforward 2D/3D anal- visualized by classical visualization methods. Such datasets are ogy, replacing the triangle of 2D ROAM by its 3D equivalent, the usually many times larger than what a video card's memory can tetrahedron. Although work in the field of hierarchical tetrahedral hold. For example, a 1024×1024×446 voxel dataset uses 446MB meshes was widely undertaken, the produced meshes were not used of memory. Rendering such datasets ”as-is” is thus not possible. for volumetric rendering purposes. We explain how to compute a Therefore, there is a need for volume visualization methods that bounded error inside the tetrahedron to build a hierarchical tetrahe- can both handle larger data sets, and have good scalability proper- dral mesh and how to refine this mesh in real time to adapt it to the ties. -

MATRIX OPERATORS and the KLEIN FOUR GROUP Ginés R Pérez Teruel
Palestine Journal of Mathematics Vol. 9(1)(2020) , 402–410 © Palestine Polytechnic University-PPU 2020 MATRIX OPERATORS AND THE KLEIN FOUR GROUP Ginés R Pérez Teruel Communicated by José Luis López Bonilla MSC 2010 Classifications: Primary 15B99,15A24; Secondary 20K99. Keywords and phrases: Klein Four Group; Per-symmetric Matrices; Matrix Algebra. I am in debt to C. Caravello for useful comments and suggestions. Abstract. In this note we show that the set of operators, S = fI; T; P; T ◦ P g that consists of the identity I, the usual transpose T , the per-transpose P and their product T ◦ P , forms a Klein Four-Group with the composition. With the introduced framework, we study in detail the properties of bisymmetric, centrosymmetric matrices and other algebraic structures, and we provide new definitions and results concerning these structures. In particular, we show that the per-tansposition allows to define a degenerate inner product of vectors, a cross product and a dyadic product of vectors with some interesting properties. In the last part of the work, we provide another realization of the Klein Group involving the tensorial product of some 2 × 2 matrices. 1 Introduction and background n×m Definition 1.1. Let A 2 R . If A = [aij] for all 1 ≤ i ≤ n, 1 ≤ j ≤ m, then P (A) is the per-transpose of A, and operation defined by P [aij] = [am−j+1;n−i+1] (1.1) Consequently, P (A) 2 Rm×n. Here, we list some properties: (i) P ◦ P (A) = A (ii) P (A ± B) = P (A) ± P (B) if A, B 2 Rn×m (iii) P (αA) = αP (A) if α 2 R (iv) P (AB) = P (B)P (A) if A 2 Rm×n, B 2 Rn×p (v) P ◦ T (A) = T ◦ P (A) where T (A) is the transpose of A (vi) det(P (A)) = det(A) (vii) P (A)−1 = P (A−1) if det(A) =6 0 The proofs of these properties follow directly from the definition. -

The Area of a Triangle
Chapter 22 The area of a triangle 22.1 Heron’s formula for the area of a triangle Theorem 22.1. The area of a triangle of sidelengths a, b, c is given by = s(s a)(s b)(s c), △ − − − 1 p where s = 2 (a + b + c). B Ia I ra r A C Y Y ′ s − b s − c s − a Proof. Consider the incircle and the excircle on the opposite side of A. From the similarity of triangles AIZ and AI′Z′, r s a = − . ra s From the similarity of triangles CIY and I′CY ′, r r =(s b)(s c). · a − − 802 The area of a triangle From these, (s a)(s b)(s c) r = − − − , r s and the area of the triangle is = rs = s(s a)(s b)(s c). △ − − − p Exercise 1. Prove that 1 2 = (2a2b2 +2b2c2 +2c2a2 a4 b4 c4). △ 16 − − − 22.2 Heron triangles 803 22.2 Heron triangles A Heron triangle is an integer triangle whose area is also an integer. 22.2.1 The perimeter of a Heron triangle is even Proposition 22.2. The semiperimeter of a Heron triangle is an integer. Proof. It is enough to consider primitive Heron triangles, those whose sides are relatively prime. Note that modulo 16, each of a4, b4, c4 is congruent to 0 or 1, according as the number is even or odd. To render in (??) the sum 2a2b2 +2b2c2 +2c2a2 a4 b4 c4 0 modulo 16, exactly two of a, b, c must be odd. It− follows− that− the≡ perimeter of a Heron triangle must be an even number. -

A Complete Bibliography of Publications in Linear Algebra and Its Applications: 2010–2019
A Complete Bibliography of Publications in Linear Algebra and its Applications: 2010{2019 Nelson H. F. Beebe University of Utah Department of Mathematics, 110 LCB 155 S 1400 E RM 233 Salt Lake City, UT 84112-0090 USA Tel: +1 801 581 5254 FAX: +1 801 581 4148 E-mail: [email protected], [email protected], [email protected] (Internet) WWW URL: http://www.math.utah.edu/~beebe/ 12 March 2021 Version 1.74 Title word cross-reference KY14, Rim12, Rud12, YHH12, vdH14]. 24 [KAAK11]. 2n − 3[BCS10,ˇ Hil13]. 2 × 2 [CGRVC13, CGSCZ10, CM14, DW11, DMS10, JK11, KJK13, MSvW12, Yan14]. (−1; 1) [AAFG12].´ (0; 1) 2 × 2 × 2 [Ber13b]. 3 [BBS12b, NP10, Ghe14a]. (2; 2; 0) [BZWL13, Bre14, CILL12, CKAC14, Fri12, [CI13, PH12]. (A; B) [PP13b]. (α, β) GOvdD14, GX12a, Kal13b, KK14, YHH12]. [HW11, HZM10]. (C; λ; µ)[dMR12].(`; m) p 3n2 − 2 2n3=2 − 3n [MR13]. [DFG10]. (H; m)[BOZ10].(κ, τ) p 3n2 − 2 2n3=23n [MR14a]. 3 × 3 [CSZ10, CR10c]. (λ, 2) [BBS12b]. (m; s; 0) [Dru14, GLZ14, Sev14]. 3 × 3 × 2 [Ber13b]. [GH13b]. (n − 3) [CGO10]. (n − 3; 2; 1) 3 × 3 × 3 [BH13b]. 4 [Ban13a, BDK11, [CCGR13]. (!) [CL12a]. (P; R)[KNS14]. BZ12b, CK13a, FP14, NSW13, Nor14]. 4 × 4 (R; S )[Tre12].−1 [LZG14]. 0 [AKZ13, σ [CJR11]. 5 Ano12-30, CGGS13, DLMZ14, Wu10a]. 1 [BH13b, CHY12, KRH14, Kol13, MW14a]. [Ano12-30, AHL+14, CGGS13, GM14, 5 × 5 [BAD09, DA10, Hil12a, Spe11]. 5 × n Kal13b, LM12, Wu10a]. 1=n [CNPP12]. [CJR11]. 6 1 <t<2 [Seo14]. 2 [AIS14, AM14, AKA13, [DK13c, DK11, DK12a, DK13b, Kar11a]. -

Volume 6 (2006) 1–16
FORUM GEOMETRICORUM A Journal on Classical Euclidean Geometry and Related Areas published by Department of Mathematical Sciences Florida Atlantic University b bbb FORUM GEOM Volume 6 2006 http://forumgeom.fau.edu ISSN 1534-1178 Editorial Board Advisors: John H. Conway Princeton, New Jersey, USA Julio Gonzalez Cabillon Montevideo, Uruguay Richard Guy Calgary, Alberta, Canada Clark Kimberling Evansville, Indiana, USA Kee Yuen Lam Vancouver, British Columbia, Canada Tsit Yuen Lam Berkeley, California, USA Fred Richman Boca Raton, Florida, USA Editor-in-chief: Paul Yiu Boca Raton, Florida, USA Editors: Clayton Dodge Orono, Maine, USA Roland Eddy St. John’s, Newfoundland, Canada Jean-Pierre Ehrmann Paris, France Chris Fisher Regina, Saskatchewan, Canada Rudolf Fritsch Munich, Germany Bernard Gibert St Etiene, France Antreas P. Hatzipolakis Athens, Greece Michael Lambrou Crete, Greece Floor van Lamoen Goes, Netherlands Fred Pui Fai Leung Singapore, Singapore Daniel B. Shapiro Columbus, Ohio, USA Steve Sigur Atlanta, Georgia, USA Man Keung Siu Hong Kong, China Peter Woo La Mirada, California, USA Technical Editors: Yuandan Lin Boca Raton, Florida, USA Aaron Meyerowitz Boca Raton, Florida, USA Xiao-Dong Zhang Boca Raton, Florida, USA Consultants: Frederick Hoffman Boca Raton, Floirda, USA Stephen Locke Boca Raton, Florida, USA Heinrich Niederhausen Boca Raton, Florida, USA Table of Contents Khoa Lu Nguyen and Juan Carlos Salazar, On the mixtilinear incircles and excircles,1 Juan Rodr´ıguez, Paula Manuel and Paulo Semi˜ao, A conic associated with the Euler line,17 Charles Thas, A note on the Droz-Farny theorem,25 Paris Pamfilos, The cyclic complex of a cyclic quadrilateral,29 Bernard Gibert, Isocubics with concurrent normals,47 Mowaffaq Hajja and Margarita Spirova, A characterization of the centroid using June Lester’s shape function,53 Christopher J. -

Trends in Field Theory Research
Complimentary Contributor Copy Complimentary Contributor Copy MATHEMATICS RESEARCH DEVELOPMENTS ADVANCES IN LINEAR ALGEBRA RESEARCH No part of this digital document may be reproduced, stored in a retrieval system or transmitted in any form or by any means. The publisher has taken reasonable care in the preparation of this digital document, but makes no expressed or implied warranty of any kind and assumes no responsibility for any errors or omissions. No liability is assumed for incidental or consequential damages in connection with or arising out of information contained herein. This digital document is sold with the clear understanding that the publisher is not engaged in rendering legal, medical or any other professional services. Complimentary Contributor Copy MATHEMATICS RESEARCH DEVELOPMENTS Additional books in this series can be found on Nova’s website under the Series tab. Additional e-books in this series can be found on Nova’s website under the e-book tab. Complimentary Contributor Copy MATHEMATICS RESEARCH DEVELOPMENTS ADVANCES IN LINEAR ALGEBRA RESEARCH IVAN KYRCHEI EDITOR New York Complimentary Contributor Copy Copyright © 2015 by Nova Science Publishers, Inc. All rights reserved. No part of this book may be reproduced, stored in a retrieval system or transmitted in any form or by any means: electronic, electrostatic, magnetic, tape, mechanical photocopying, recording or otherwise without the written permission of the Publisher. For permission to use material from this book please contact us: [email protected] NOTICE TO THE READER The Publisher has taken reasonable care in the preparation of this book, but makes no expressed or implied warranty of any kind and assumes no responsibility for any errors or omissions. -

On Triangles As Real Analytic Varieties of an Extended Fermat Equation
On Triangles as Real Analytic Varieties of an Extended Fermat Equation Giri Prabhakar∗ Abstract On the one hand, triangles play an important role in the solution of elliptic curves, as exemplified in studies by Tunnell on the congruent number problem, or Ono relating arbitrary triangles to elliptic curves. On the other hand, Frey-Hellegouarch curves are deeply connected to Fermat's equation. Thus, we are motivated to explore the relationship between triangles and Fermat's equation. We show that triangles in Euclidean space can be exactly described by real analytic varieties of an extension of the Fermat equation. To the 3 best of our knowledge, this formulation has not been explored previously. Let S = f(a; b; c) 2 R≥0 j (c > φ φ φ 0 ^ 9 φ 2 R≥1)[a + b = c ]g be the set of real non-negative solutions to the Fermat equation extended to admit real valued exponents φ ≥ 1, which we term the Fermat index. Let E be the set of triplets of side lengths of all non-degenerate and degenerate 2-simplices. We show that S = E, and that each triangle has a unique Fermat index. We show that there exists a deformation retraction F based on the extended Fermat equation, that maps all possible sides a ≤ c and b ≤ c of 2-simplices (a; b; c) 2 S, to the retract c of fixed length at a fixed angle to a. However, (a; b; c) is also governed by the law of cosines; thus an alternate map P exists based on the law of cosines, which must be geometrically identical to F . -

Integer Trigonometry for Integer Angles
Algorithms and Computation in Mathematics • Volume 26 Editors Arjeh M. Cohen Henri Cohen David Eisenbud Michael F. Singer Bernd Sturmfels For further volumes: http://www.springer.com/series/3339 Oleg Karpenkov Geometry of Continued Fractions Oleg Karpenkov Dept. of Mathematical Sciences University of Liverpool Liverpool, UK ISSN 1431-1550 Algorithms and Computation in Mathematics ISBN 978-3-642-39367-9 ISBN 978-3-642-39368-6 (eBook) DOI 10.1007/978-3-642-39368-6 Springer Heidelberg New York Dordrecht London Library of Congress Control Number: 2013946250 Mathematics Subject Classification (2010): 11J70, 11H06, 11P21, 52C05, 52C07 © Springer-Verlag Berlin Heidelberg 2013 This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. Exempted from this legal reservation are brief excerpts in connection with reviews or scholarly analysis or material supplied specifically for the purpose of being entered and executed on a computer system, for exclusive use by the purchaser of the work. Duplication of this publication or parts thereof is permitted only under the provisions of the Copyright Law of the Publisher’s location, in its current version, and permission for use must always be obtained from Springer. Permissions for use may be obtained through RightsLink at the Copyright Clearance Center. Violations are liable to prosecution under the respective Copyright Law. -

The General Coupled Matrix Equations Over Generalized Bisymmetric Matrices ∗ Mehdi Dehghan , Masoud Hajarian
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Linear Algebra and its Applications 432 (2010) 1531–1552 Contents lists available at ScienceDirect Linear Algebra and its Applications journal homepage: www.elsevier.com/locate/laa The general coupled matrix equations over generalized bisymmetric matrices ∗ Mehdi Dehghan , Masoud Hajarian Department of Applied Mathematics, Faculty of Mathematics and Computer Science, Amirkabir University of Technology, No. 424, Hafez Avenue, Tehran 15914, Iran ARTICLE INFO ABSTRACT Article history: Received 20 May 2009 In the present paper, by extending the idea of conjugate gradient Accepted 12 November 2009 (CG) method, we construct an iterative method to solve the general Available online 16 December 2009 coupled matrix equations Submitted by S.M. Rump p AijXjBij = Mi,i= 1, 2, ...,p, j=1 AMS classification: 15A24 (including the generalized (coupled) Lyapunov and Sylvester matrix 65F10 equations as special cases) over generalized bisymmetric matrix 65F30 group (X1,X2, ...,Xp). By using the iterative method, the solvability of the general coupled matrix equations over generalized bisym- Keywords: metric matrix group can be determined in the absence of roundoff Iterative method errors. When the general coupled matrix equations are consistent Least Frobenius norm solution group over generalized bisymmetric matrices, a generalized bisymmetric Optimal approximation generalized solution group can be obtained within finite iteration steps in the bisymmetric solution group Generalized bisymmetric matrix absence of roundoff errors. The least Frobenius norm generalized General coupled matrix equations bisymmetric solution group of the general coupled matrix equa- tions can be derived when an appropriate initial iterative matrix group is chosen. -

A. Appendix: Some Mathematical Concepts
A. APPENDIX: SOME MATHEMATICAL CONCEPTS Overview This appendix on mathematical concepts is simply intended to aid readers who would benefit from a refresher on material they learned in the past or possibly never learned at all. Included here are basic definitions and examples pertaining to aspects of set theory, numerical linear algebra, met- ric properties of sets in real n-space, and analytical properties of functions. Among the latter are such matters as continuity, differentiability, and con- vexity. A.1 Basic terminology and notation from set theory This section recalls some general definitions and notation from set theory. In Section A.4 we move on to consider particular kinds of sets; in the process, we necessarily draw certain functions into the discussion. A set is a collection of objects belonging to some universe of discourse. The objects belonging to the collection (i.e., set) are said to be its members or elements.Thesymbol∈ is used to express the membership of an object in a set. If x is an object and A is a set, the statement that x is an element of A (or x belongs to A) is written x ∈ A. To negate the membership of x in the set A we write x/∈ A. The set of all objects, in the universe of discourse, not belonging to A is called the complement of A. There are many different notations used for the complement of a set A. One of the most popular is Ac. The cardinality of a set is the number of its elements. A set A can have cardinality zero in which case it is said to be empty. -

PROBLEMS in ELEMENTARY NUMBER THEORY Hojoo
PROBLEMS IN ELEMENTARY NUMBER THEORY Hojoo Lee Version 050722 God does arithmetic. C. F. Gauss 1, −24, 252, −1472, 4830, −6048, −16744, 84480, −113643, −115920, 534612, −370944, −577738, 401856, 1217160, 987136, −6905934, 2727432, 10661420, −7109760, −4219488, −12830688, 18643272, 21288960, −25499225, 13865712, −73279080, 24647168, ··· 1 2 PROBLEMS IN ELEMENTARY NUMBER THEORY Contents 1. Introduction 3 2. Notations and Abbreviations 4 3. Divisibility Theory I 5 4. Divisibility Theory II 12 5. Arithmetic in Zn 16 Primitive Roots 16 Quadratic Residues 17 Congruences 17 6. Primes and Composite Numbers 20 Composite Numbers 20 Prime Numbers 20 7. Rational and Irrational Numbers 24 Rational Numbers 24 Irrational Numbers 25 8. Diophantine Equations I 29 9. Diophantine Equations II 34 10. Functions in Number Theory 37 Floor Function and Fractional Part Function 37 Euler phi Function 39 Divisor Functions 39 More Functions 40 Functional Equations 41 11. Polynomials 44 12. Sequences of Integers 46 Linear Recurrnces 46 Recursive Sequences 47 More Sequences 51 13. Combinatorial Number Theory 54 14. Additive Number Theory 61 15. The Geometry of Numbers 66 16. Miscellaneous Problems 67 17. Sources 71 18. References 94 PROBLEMS IN ELEMENTARY NUMBER THEORY 3 1. Introduction The heart of Mathematics is its problems. Paul Halmos 1. Introduction Number Theory is a beautiful branch of Mathematics. The purpose of this book is to present a collection of interesting questions in Number Theory. Many of the problems are mathematical competition problems all over the world including IMO, APMO, APMC, and Putnam, etc. The book is available at http://my.netian.com/∼ideahitme/orange.html 2. -

Two Brahmagupta Problems
Forum Geometricorum b Volume 6 (2006) 301–310. bbb FORUM GEOM ISSN 1534-1178 Two Brahmagupta Problems K. R. S. Sastry Abstract. D. E. Smith reproduces two problems from Brahmagupta’s work Ku- takh¯adyaka (algebra) in his History of Mathematics, Volume 1. One of them involves a broken tree and the other a mountain journey. Normally such objects are represented by vertical line segments. However, it is every day experience that such objects need not be vertical. In this paper, we generalize these situations to include slanted positions and provide integer solutions to these problems. 1. Introduction School textbooks on geometry and trigonometry contain problems about trees, poles, buildings, hills etc. to be solved using the Pythagorean theorem or trigono- metric ratios. The assumption is that such objects are vertical. However, trees grow not only vertically (and tall offering a magestic look) but also assume slanted positions (thereby offering an elegant look). Buildings too need not be vertical in structure, for example the leaning tower of Pisa. Also, a distant planar view of a mountain is more like a scalene triangle than a right one. In this paper we regard the angles formed in such situations as having rational cosines. We solve the fol- lowing Brahmagupta problems from [5] in the context of rational cosines triangles. In [4] these problems have been given Pythagorean solutions. Problem 1. A bamboo 18 cubits high was broken by the wind. Its tip touched the ground 6 cubits from the root. Tell the lengths of the segments of the bamboo. Problem 2. On the top of a certain hill live two ascetics.