Protozoology from the Perspective of Science Theory: History and Concept of a Biological Discipline
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

On the Development of Spinoza's Account of Human Religion
Intermountain West Journal of Religious Studies Volume 5 Number 1 Spring 2014 Article 4 2014 On the Development of Spinoza’s Account of Human Religion James Simkins University of Pittsburgh Follow this and additional works at: https://digitalcommons.usu.edu/imwjournal Recommended Citation Simkins, James "On the Development of Spinoza’s Account of Human Religion." Intermountain West Journal of Religious Studies 5, no. 1 (2014). https://digitalcommons.usu.edu/imwjournal/ vol5/iss1/4 This Article is brought to you for free and open access by the Journals at DigitalCommons@USU. It has been accepted for inclusion in Intermountain West Journal of Religious Studies by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. 52 James Simkins: On the Development of Spinoza’s Account of Human Religion JAMES SIMKINS graduated with philosophy, history, and history and philosophy of science majors from the University of Pittsburgh in 2013. He is currently taking an indefinite amount of time off to explore himself and contemplate whether or not to pursue graduate study. His academic interests include Spinoza, epistemology, and history from below. IMW Journal of Religious Studies Vol. 5:1 53 ‡ On the Development of Spinoza’s Account of Human Religion ‡ In his philosophical and political writings, Benedict Spinoza (1632-1677) develops an account of human religion, which represents a unique theoretical orientation in the early modern period.1 This position is implicit in many of Spinoza’s philosophical arguments in the Treatise on the Emendation of the Intellect, the Short Treatise, and Ethics.2 However, it is most carefully developed in his Tractatus Theologico-Politicus (hereafter TTP).3 What makes Spinoza’s position unique is the fact that he rejects a traditional conception of religion on naturalistic grounds, while refusing to dismiss all religion as an entirely anthropological phenomenon. -

Menasseh Ben Israel and His World Brill's Studies in Intellectual History
MENASSEH BEN ISRAEL AND HIS WORLD BRILL'S STUDIES IN INTELLECTUAL HISTORY General Editor AJ. VANDERJAGT, University of Groningen Editorial Board M. COLISH, Oberlin College J.I. ISRAEL, University College, London J.D. NORTH, University of Groningen R.H. POPKIN, Washington University, St. Louis-UCLA VOLUME 15 MENASSEH BEN ISRAEL AND HIS WORLD EDITED BY YOSEF KAPLAN, HENRY MECHOULAN AND RICHARD H. POPKIN ^o fr-hw'* -A EJ. BRILL LEIDEN • NEW YORK • K0BENHAVN • KÖLN 1989 Published with financial support from the Dr. C. Louise Thijssen- Schoutestichting. Library of Congress Cataloging-in-Publication Data Menasseh Ben Israel and his world / edited by Yosef Kaplan, Henry Méchoulan and Richard H. Popkin. p. cm. -- (Brill's studies in intellectual history, ISSN 0920-8607 ; v. 15) Includes index. ISBN 9004091149 1. Menasseh ben Israel, 1604-1657. 2. Rabbis-Netherlands- -Amsterdam-Biography. 3. Amsterdam (Netherlands)-Biography. 4. Sephardim--Netherlands--Amsterdam--History--17th century. 5. Judaism--Netherlands--Amsterdam--History--17th century. I. Kaplan, Yosef. II. Popkin, Richard Henry, 1923- BM755.M25M46 1989 296'.092-dc20 89-7265 [B] CIP ISSN 0920-8607 ISBN 90 04 09114 9 © Copyright 1989 by E.J. Brill, The Netherlands All rights reserved. No part of this book may be reproduced or translated in any form, by print, photoprint, microfilm, microfiche or any other means without written permission from the publisher PRINTED IN THE NETHERLANDS BY E.J. BRILI, CONTENTS Introduction, Richard H. Popkin vu A Generation of Progress in the Historical Study of Dutch Sephardic Jewry, Yosef Kaplan 1 The Jewish Dimension of the Scottish Apocalypse: Climate, Cove- nant and World Renewal, Arthur H. -

The Intestinal Protozoa
The Intestinal Protozoa A. Introduction 1. The Phylum Protozoa is classified into four major subdivisions according to the methods of locomotion and reproduction. a. The amoebae (Superclass Sarcodina, Class Rhizopodea move by means of pseudopodia and reproduce exclusively by asexual binary division. b. The flagellates (Superclass Mastigophora, Class Zoomasitgophorea) typically move by long, whiplike flagella and reproduce by binary fission. c. The ciliates (Subphylum Ciliophora, Class Ciliata) are propelled by rows of cilia that beat with a synchronized wavelike motion. d. The sporozoans (Subphylum Sporozoa) lack specialized organelles of motility but have a unique type of life cycle, alternating between sexual and asexual reproductive cycles (alternation of generations). e. Number of species - there are about 45,000 protozoan species; around 8000 are parasitic, and around 25 species are important to humans. 2. Diagnosis - must learn to differentiate between the harmless and the medically important. This is most often based upon the morphology of respective organisms. 3. Transmission - mostly person-to-person, via fecal-oral route; fecally contaminated food or water important (organisms remain viable for around 30 days in cool moist environment with few bacteria; other means of transmission include sexual, insects, animals (zoonoses). B. Structures 1. trophozoite - the motile vegetative stage; multiplies via binary fission; colonizes host. 2. cyst - the inactive, non-motile, infective stage; survives the environment due to the presence of a cyst wall. 3. nuclear structure - important in the identification of organisms and species differentiation. 4. diagnostic features a. size - helpful in identifying organisms; must have calibrated objectives on the microscope in order to measure accurately. -

The Life Cycle of Trypanosoma Cruzi
Tyler et al. THE LIFE CYCLE OF TRYPANOSOMA CRUZI K. M. Tyler, C. L. Olson and D. M. Engman Departments of Microbiology-Immunology and Pathology Feinberg Medical School of Northwestern University, Chicago, IL 60611 ABSTRACT Since the discovery of Trypanosoma cruzi as the parasite that causes Chagas disease, nearly a century ago, the details of the organism's life cycle have fascinated scientists. T. cruzi is a single-celled eukaryote with a complex life cycle alternating between reduviid bug vectors and vertebrate hosts. It is able to adapt via the process of cellular differentiation to replicate within the diverse environments represented of the insect's gut and host cell cytoplasm. These adaptive transformations take the form of coordinated changes in morphology, metabolism and cell cycle regulation. Different life cycle stages of T. cruzi show dramatically different protein and RNA profiles, which are the end result of unusual mechanisms for regulating gene expression. In recent years, new molecular techniques have been brought to bear on the life cycle dramatically increasing our knowledge of the strategies employed by the parasite to ensure its continued survival. INTRODUCTION Chagas disease The etiologic agent of the chronic and often fatal Chagas disease is the American trypanosome, Trypanosoma cruzi , a flagellated protozoan of the order Kinetoplastida. The survival of T. cruzi is dependent on the successful transmission between, and the colonization of, two radically different environments: the midgut of the reduviid bug vector and the cytoplasm of the mammalian host cell. As is true of all infections, interruption of the pathogen's life cycle will lead to eradication of the disease. -

A PRELIMINARY NOTE on TETRAMITUS, a STAGE in the LIFE CYCLE of a COPROZOIC AMOEBA by MARTHA BUNTING
294 Z6OL6G Y.- M. BUNTING PROC. N. A. S. A PRELIMINARY NOTE ON TETRAMITUS, A STAGE IN THE LIFE CYCLE OF A COPROZOIC AMOEBA By MARTHA BUNTING DEPARTMENT OF ZOOLOGY, UNIVERSITY OF PENNSYLVANIA Communicated July 24, 1922 In 1920 a study of the Entamoeba of the rat was begun and in a culture on artificial medium a number of Coprozoic amoeba appeared, one of which exhibited a flagellate phase which seems to be identical with Tetramitus rostratus Perty.1 The appearance of flagellate phases in cultures of Coprozoic amoeba (Whitmore,2 etc.) as well as of soil amoeba (Kofoid,1 Wilson,4 etc.) has been recorded a number of times, but the flagellates which appeared were relatively simple in organization and very transitory in occurrence. Fur- thermore, some of the simpler flagellates have been observed (Pascher5) to lose their flagella and become amoeboid. In the case here described, however, there is a transformation of a simple amoeba into a relatively complex flagellate which multiplies for several days then returns to the amoeboid condition and becomes encysted. Tetramitus rostratus has been studied by a number of investigators since its discovery in 1852, by Perty,' yet no one has given a full account of its life history. The lack of cysts in previous accounts, mentioned by Dobell and O'Connor,6 is explained by the transformation into an amoeba before encystment. It is believed that this animal presents the extreme in transformations of this sort and serves to emphasize the close relationship between amoebae and flagellates, and the need for careful studies of life cycles, in pure cultures where possible, in investigations of coprozoic and other Protozoa. -

What Is a Microorganism?
Microbiology: What is it? ! Study of organisms who are too small to be seen without a microscope. ! Study of small organisms or microorganisms. NOT just Bacteria! ! Study of single celled organisms. The original cell biology! ! Categories & subjects based on the type of organisms: (1) Viruses – Virology (acellular) (2a) Bacteria – Bacteriology (e.g. Prokaryotes) (2b) Archea – Archeaology? (already taken) (3) Fungi – Mycology (4) Algae – Phycology (5) Protozoa – Protozoology WHAT IS A MICROORGANISM? “There is no simple answer to this question. The word ‘microorganism’ is not the name of a group of related organisms, as are the words ‘plants’ or ‘invertebrates’ or ‘fish’. The use of the word does, however, indicate that there is something special about small organisms; we use no special word to denote large organisms or medium-sized ones. - Sistrom (1969) 1 Reasons to study Microbiology: (1) Bacteria are part of us! E. coli lives in our gut and produces essential vitamins (e.g. K). (2) Infectivity & Pathogenicity; MO’s have the ability to cause disease in compromised &/or heathy hosts. (3) MO’s in the environment; Bioremediation or use of MO’s to breakdown waste compounds like oil, pesticides, etc. Mineral cycling of elements like N, S, Fe, etc. (4) Applied Microbiology or use in agriculture and industry. (5) Understand basic biological processes: Evolution, Ecology, Genetics, etc. WHY STUDY MICROBIOLOGY? “The role of the infinitely small is infinitely large.” - Louis Pasteur (1862) 2 WE ARE NOT ALONE! “We are outnumbered. The average human contains about 10 trillion cells. On that average human are about 10 times as many microorganisms, or 100 trillion cells...As long as they stay in balance and where they belong, [they] do us no harm…In fact, many of them provide some important services to us. -

A Monograph on the Protozoa of the Large Intestine of the Horse Ta-Shih Hsiung Iowa State College
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1930 A monograph on the protozoa of the large intestine of the horse Ta-Shih Hsiung Iowa State College Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Zoology Commons Recommended Citation Hsiung, Ta-Shih, "A monograph on the protozoa of the large intestine of the horse" (1930). Retrospective Theses and Dissertations. 14768. https://lib.dr.iastate.edu/rtd/14768 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overiaps. -

Baruch Spinoza Chronology
Baruch Spinoza Chronology 1391 Spanish Jews are forced to convert to Catholicism for the sake of "social and sectarian uniformity." 1478 Establishment of the Spanish Inquisition, whose primary task is to convict and execute those found "judaizing." 1492 All practising Jews in Spain are given the choice to convert or be expelled. 1497 All Portuguese Jews (including Spinoza’s ancestors) are forced to convert. A steady stream of Jewish refugees begins to flow from Portugal. 1587/8 Spinoza’s father Michael is born in Vidigere, Portugal, to Isaac d’Espinoza 1609 Beginning of the twelve year truce between the United Provinces and Spain, effectively establishing political independence (after nearly a 100 year struggle) for the seven northern provinces as well as their (Protestant) sectarian separation from the (Catholic) southern provinces. 1618 Defenestration of Prague and beginning of the Thirty Years War. Calvinist-inspired coup d’état in the Dutch Republic, led by the Prince of Orange, leading to the execution of Oldenbarnevelt and imprisonment of Grotius. Uriel d’Acosta (or da Costa), a Portuguese “New Christian” who had returned to Judaism in Amsterdam but became disillusioned with the Jewish community, is excommunicated for the first time in Venice for denying the immortality of the soul and questioning the Mosaic authorship of the Torah, a decree later affirmed in Amsterdam in 1623 and renewed in 1633. 1619 Batavia, Java is established as headquarters of the Dutch East India Company. 1620 Francis Bacon writes Novum organum. 1621 Hostilities resume between Spain and the United Provinces. 1622 Probable date Spinoza’s father arrives in Amsterdam, probably from Nantes. -
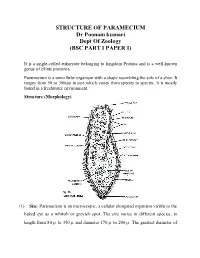
STRUCTURE of PARAMECIUM Dr Poonam Kumari Dept of Zoology (BSC PART I PAPER I)
STRUCTURE OF PARAMECIUM Dr Poonam kumari Dept Of Zoology (BSC PART I PAPER I) It is a single-celled eukaryote belonging to kingdom Protista and is a well-known genus of ciliate protozoa. Paramecium is a unicellular organism with a shape resembling the sole of a shoe. It ranges from 50 to 300um in size which varies from species to species. It is mostly found in a freshwater environment. Structure (Morphology) (1) Size: Paramecium is an microscopic, a cellular elongated organism visible to the baked eye as a whitish or greyish spot. The size varies in different species, in length from 80 to 350 and diameter 170 to 290 . The greatest diameter of the cylindrical body is about two third of its entire length. Usually the individuals of the same species may show minor morphological and physiological differences. (2) Shape: Paramecium is a slipper shaped, cigar shaped, or spindle shaped animalcule. Its shape is usually constant and a symmetrical, because slipper like shape. The body is elongated, blunt and rounded at the anterior end and somewhat pointed of the posterior end. In cross section it is circular with greatest diameter behind the centre of body. The anterior half of the body is slightly twisted. The body is distinguished into an oral or ventral surface and an aboral or dorsal surface. The structure is more complicated due to the development of certain organelles in the acellular body. (3) Oral groove: The ventral surface of body bears a prominent, oblique and shallow depression is called oral groove, it arise from the middle of body and extends to the left side of anterior end. -

Menasseh Ben Israel's Christian Connection: Henry Jessey and the Jews
MENASSEH BEN ISRAEL'S CHRISTIAN CONNECTION: HENRY JESSEY AND THE JEWS DAVID S. KATZ "He was of a middle Stature, and inclining to Fatness", an English con temporary described Rabbi Menasseh ben Israel, "He always wore his own Hair, which (many years before his Death) was very Grey; so that his Complexion being pretty fresh, his Demeanor Grace ful, and Comely, his Habit plain and decent, he Commanded an aweful Rev erence which was justly due to so venerable a Deportment: In short, he was un homme sans Passion, sans légèreté, mais Hélas/ sans opulence' '1. From his lodgings in the Strand, Menasseh sallied forth to meet politi cians, divines, intellectuals, and anyone who conceivably could help him to reach the goal of his English mission: an official authorization of the re- admission of the Jews to England after an exile of over three and a half centuries2. Menasseh had come to London in September 1655, and al though the Whitehall Conference of December failed to resolve the re- admission question, he remained in England for exactly two years in the vain hope of obtaining a formal written permission3. During his stay in London, he seems to have established himself, at least among gentiles, as a self-appointed ambassador of world Jewry and as a renowned expert in things Jewish. Menasseh received Ralph Cudworth, the Regius professor of Hebrew, and gave him a manuscript summarizing the Jewish objections to Christianity4. He discussed plans for a polyglot bible with Henry Thorndike5. He held further meetings with Henry Ol denburg, later secretary of the Royal Society; with Adam Boreel, the Con- 1 Menasseh ben Israel, Of The Term of Liß, ed. -

Lamarck: the Birth of Biology Author(S): Frans A
Lamarck: The Birth of Biology Author(s): Frans A. Stafleu Reviewed work(s): Source: Taxon, Vol. 20, No. 4 (Aug., 1971), pp. 397-442 Published by: International Association for Plant Taxonomy (IAPT) Stable URL: http://www.jstor.org/stable/1218244 . Accessed: 24/12/2012 16:29 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. International Association for Plant Taxonomy (IAPT) is collaborating with JSTOR to digitize, preserve and extend access to Taxon. http://www.jstor.org This content downloaded on Mon, 24 Dec 2012 16:29:36 PM All use subject to JSTOR Terms and Conditions TAXON 20(4): 397-442. AUGUST 1971 LAMARCK:THE BIRTH OF BIOLOGY Frans A. Stafleu "A long blind patience, such was his genius of the Universe" (Sainte Beuve) Summary A review of the development of Lamarck'sideas on biological systematibswith special reference to the origin and development of his concept of organic evolution. Lamarck's development towards biological systematics is traced through his early botanical and geological writings and related to the gradual change in his scientific outlook from a static and essentialist view of nature towards a dynamic and positivist concept of the life sciences as a special discipline. -

Jean-Baptiste Lamarck, Philosophie Zoologique
Jean-Baptiste Lamarck, Philosophie Zoologique (1809) Chapter 7: Concerning the Influence of Circumstances on the Actions and Habits of Animals, and the Influence of the Actions and Habits of these Living Bodies As Causes Which Modify Their Organic Structure and Their Parts What we are now concerned with is not a rational speculation but the examination of a reliable fact, a more universal one than people think and something to which we have neglected to pay the attention it deserves. Undoubtedly this is the case because on most occasions it is very difficult to recognize. This fact consists of the influence which circumstances exert on the different living things subject to them. In truth, for quite a long time now we have noticed the influence of the different states of our organic structure on our characteristics, inclinations, actions, and even our ideas. But it seems to me that no one has get made known the influence of our actions and habits on our own organic structure. Now, as these actions and habits are entirely dependent on the circumstances in which we usually find ourselves, I am going to try to show how great the influence is which these circumstances exert on the general form, the condition of the parts, and even on the organic structure of living things. Thus, this chapter is going to explore this very well established fact. If we had not had numerous occasions to recognize quite clearly the effects of this influence on certain living bodies which we have transported into entirely new environments, very different from the ones where they used to live, and if we had not seen these effects and the changes resulting from them come to light in some way under our very eyes, the important fact under discussion would have always remained unknown to us.