Mekonnen Et Al 2012
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
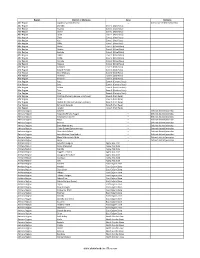
Districts of Ethiopia
Region District or Woredas Zone Remarks Afar Region Argobba Special Woreda -- Independent district/woredas Afar Region Afambo Zone 1 (Awsi Rasu) Afar Region Asayita Zone 1 (Awsi Rasu) Afar Region Chifra Zone 1 (Awsi Rasu) Afar Region Dubti Zone 1 (Awsi Rasu) Afar Region Elidar Zone 1 (Awsi Rasu) Afar Region Kori Zone 1 (Awsi Rasu) Afar Region Mille Zone 1 (Awsi Rasu) Afar Region Abala Zone 2 (Kilbet Rasu) Afar Region Afdera Zone 2 (Kilbet Rasu) Afar Region Berhale Zone 2 (Kilbet Rasu) Afar Region Dallol Zone 2 (Kilbet Rasu) Afar Region Erebti Zone 2 (Kilbet Rasu) Afar Region Koneba Zone 2 (Kilbet Rasu) Afar Region Megale Zone 2 (Kilbet Rasu) Afar Region Amibara Zone 3 (Gabi Rasu) Afar Region Awash Fentale Zone 3 (Gabi Rasu) Afar Region Bure Mudaytu Zone 3 (Gabi Rasu) Afar Region Dulecha Zone 3 (Gabi Rasu) Afar Region Gewane Zone 3 (Gabi Rasu) Afar Region Aura Zone 4 (Fantena Rasu) Afar Region Ewa Zone 4 (Fantena Rasu) Afar Region Gulina Zone 4 (Fantena Rasu) Afar Region Teru Zone 4 (Fantena Rasu) Afar Region Yalo Zone 4 (Fantena Rasu) Afar Region Dalifage (formerly known as Artuma) Zone 5 (Hari Rasu) Afar Region Dewe Zone 5 (Hari Rasu) Afar Region Hadele Ele (formerly known as Fursi) Zone 5 (Hari Rasu) Afar Region Simurobi Gele'alo Zone 5 (Hari Rasu) Afar Region Telalak Zone 5 (Hari Rasu) Amhara Region Achefer -- Defunct district/woredas Amhara Region Angolalla Terana Asagirt -- Defunct district/woredas Amhara Region Artuma Fursina Jile -- Defunct district/woredas Amhara Region Banja -- Defunct district/woredas Amhara Region Belessa -- -

Nigella Sativa) at the Oromia Regional State, Ethiopia
Asian Journal of Agricultural Extension, Economics & Sociology 31(3): 1-12, 2019; Article no.AJAEES.47315 ISSN: 2320-7027 Assessment of Production and Utilization of Black Cumin (Nigella sativa) at the Oromia Regional State, Ethiopia Wubeshet Teshome1 and Dessalegn Anshiso2* 1Ethiopian Biodiversity Institute, Horticulture and Crop Biodiversity Directorate, P.O.Box 30726; Addis Ababa, Ethiopia. 2College of Economics and Management, Huazhong Agricultural University, No. 1 Shizishan Street, Hongshan District, Wuhan, 430070, Hubei, P.R. China. Authors’ contributions This work was carried out in collaboration between both authors. Author WT managed the literature searches and participated in data collection. Author DA designed the study, performed the statistical analysis, wrote the protocol and wrote the first draft of the manuscript. Both authors read and approved the final manuscript. Article Information DOI: 10.9734/AJAEES/2019/v31i330132 Editor(s): (1) Prof. Fotios Chatzitheodoridis, Department of Agricultural Technology-Division of Agricultural Economics, Technological Education Institute of Western Macedonia, Greece. Reviewers: (1) Lawal Mohammad Anka, Development Project Samaru Gusau Zamfara State, Nigeria. (2) İsmail Ukav, Adiyaman University, Turkey. Complete Peer review History: http://www.sdiarticle3.com/review-history/47315 Received 14 November 2018 Accepted 09 February 2019 Original Research Article Published 06 April 2019 ABSTRACT Background and Objective: Black cuminseed for local consumption and other importance, such as oil and oil rosin for medicinal purposes, export market, crop diversification, income generation, reducing the risk of crop failure and others made it as a best alternative crop under Ethiopian smaller land holdings. The objectives of this study were to examine factors affecting farmer perception of the Black cumin production importance, and assess the crop utilization purpose by smallholder farmers and its income potential for the farmers in two Districts of Bale zone of Oromia regional state in Ethiopia. -

Body Measurements for the Monkeys of Bioko Island, Equatorial Guinea
Primate Conservation 2009 (24): 99–105 Body Measurements for the Monkeys of Bioko Island, Equatorial Guinea Thomas M. Butynski¹,², Yvonne A. de Jong² and Gail W. Hearn¹ ¹Bioko Biodiversity Protection Program, Drexel University, Philadelphia, PA, USA ²Eastern Africa Primate Diversity and Conservation Program, Nanyuki, Kenya Abstract: Bioko Island, Equatorial Guinea, has a rich (eight genera, 11 species), unique (seven endemic subspecies), and threat- ened (five species) primate fauna, but the taxonomic status of most forms is not clear. This uncertainty is a serious problem for the setting of priorities for the conservation of Bioko’s (and the region’s) primates. Some of the questions related to the taxonomic status of Bioko’s primates can be resolved through the statistical comparison of data on their body measurements with those of their counterparts on the African mainland. Data for such comparisons are, however, lacking. This note presents the first large set of body measurement data for each of the seven species of monkeys endemic to Bioko; means, ranges, standard deviations and sample sizes for seven body measurements. These 49 data sets derive from 544 fresh adult specimens (235 adult males and 309 adult females) collected by shotgun hunters for sale in the bushmeat market in Malabo. Key Words: Bioko Island, body measurements, conservation, monkeys, morphology, taxonomy Introduction gordonorum), and surprisingly few such data exist even for some of the more widespread species (for example, Allen’s Comparing external body measurements for adult indi- swamp monkey Allenopithecus nigroviridis, northern tala- viduals from different sites has long been used as a tool for poin monkey Miopithecus ogouensis, and grivet Chlorocebus describing populations, subspecies, and species of animals aethiops). -

World Bank Document
PROCUREMENT PLAN (Textual Part) Project information: Ethiopia, One WASH- Consolidated WASH Account (CWA) Project “Phase II”, P167794 Project Implementation agency: Water Development Commission (WDC) of Public Disclosure Authorized the Ministry of Water, Irrigation and Energy (MoWIE) Date of the Procurement Plan: August 28, 2019 Period covered by this Procurement Plan: September 2019 to August 2020. Preamble In accordance with paragraph 5.9 of the “World Bank Procurement Regulations for IPF Borrowers” (July 2016 revised August 2018) (“Procurement Regulations”) the Bank’s Systematic Tracking and Exchanges in Procurement (STEP) system will be used to prepare, clear and update Procurement Plans and conduct all procurement Public Disclosure Authorized transactions for the Project. This textual part along with the Procurement Plan tables in STEP constitute the Procurement Plan for the Project. The following conditions apply to all procurement activities in the Procurement Plan. The other elements of the Procurement Plan as required under paragraph 4.4 of the Procurement Regulations are set forth in STEP. The Bank’s Standard Procurement Documents: shall be used for all contracts subject to international competitive procurement and those contracts as specified in the Procurement Plan tables in STEP. Public Disclosure Authorized National Procurement Arrangements: In accordance with paragraph 5.3 of the Procurement Regulations, when approaching the national market (as specified in the Procurement Plan tables in STEP), the country’s own procurement procedures may be used. When the Borrower uses its own national open competitive procurement arrangements as set forth in Section 33(1)(a) from 35-48 of the Proclamation Number 649/2009 of the Ethiopian Federal Government Procurement and Property Administration Proclamation, such arrangements shall be subject to paragraph 5.4 of the Procurement Regulations and the following conditions. -

Determinants and Role of Farmers' Seed and Seedling
DETERMINANTS AND ROLE OF FARMERS’ SEED AND SEEDLING MULTIPLICATION IN THE SNNP REGION SEED SYSTEM M.Sc. Thesis GEZAHAGN WALELIGN April 2008 Haramaya University DETERMINANTS AND ROLE OF FARMERS’ SEED AND SEEDLING MULTIPLICATION IN THE SNNP REGION SEED SYSTEM A Thesis Submitted to the Department of Agricultural Economics, School of Graduate Studies Haramaya University In Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE IN AGRICULTURE (AGRICULTURAL ECONOMICS) By GEZAHAGN WALELIGN April 2008 Haramaya University ii School of Graduate Studies Haramaya University As Thesis Research advisor, I here by certify that I have read and evaluated this thesis prepared, under my guidance, by Gezahagn Walelign, entitled Determinants and Role of Farmers’ Seed and Seedling Multiplication in the SNNP Regional Seed System. I recommend that it be submitted as fulfillment of the Thesis requirement. Dawit Alemu(Ph D) ________________ _______________ Major Advisor Signature Date R.S.L.Srivastava (prof.) _________________ _______________ Co-advisor Signature Date As member of the Board of Examiners of the MSc Thesis Open Defense Examination, We certify that we have read, evaluated the Thesis prepared by Gezahagn Walelign and examined the candidate. We recommended that the Thesis is accepted as fulfilling the Thesis requirement for the Degree of Master of Science in Agriculture (Agricultural Economics). ______________________ _________________ _______________ Chairperson Signature Date ______________________ _________________ _______________ Internal Examiner Signature Date ______________________ _________________ _______________ External Examiner Signature Date iii DEDICATION I dedicated this thesis manuscript to my late wife Serkalem Chalachew whom I lost during my stay here for MSc. course and my beloved children Ruth and Tewodrose Gezahagn iv STATEMENT OF AUTHOR First, I declare that this thesis is the result of my own work and that all sources or materials used for this thesis have been duly acknowledged. -

Hum Ethio Manitar Opia Rian Re Espons E Fund D
Hum anitarian Response Fund Ethiopia OCHA, 2011 OCHA, 2011 Annual Report 2011 Office for the Coordination of Humanitarian Affairs Humanitarian Response Fund – Ethiopia Annual Report 2011 Table of Contents Note from the Humanitarian Coordinator ................................................................................................ 2 Acknowledgements ................................................................................................................................. 3 1. Executive Summary ............................................................................................................................ 4 1.1 2011 Humanitarian Context ........................................................................................................... 4 1.2 Map - 2011 HRF Supported Projects ............................................................................................. 6 2. Information on Contributors ................................................................................................................ 7 2.1 Donor Contributions to HRF .......................................................................................................... 7 3. Fund Overview .................................................................................................................................... 8 3.1 Summary of HRF Allocations in 2011 ............................................................................................ 8 3.1.1 HRF Allocation by Sector ....................................................................................................... -

Low Paternity Skew and the Influence of Maternal Kin in an Egalitarian
Low paternity skew and the influence of maternal kin in an egalitarian, patrilocal primate Karen B. Striera,1, Paulo B. Chavesb, Sérgio L. Mendesc, Valéria Fagundesc, and Anthony Di Fioreb,d aDepartment of Anthropology, University of Wisconsin, Madison, WI 53706; bDepartment of Anthropology and Center for the Study of Human Origins, New York University, New York, NY 10003; cDepartamento de Ciências Biológicas, Centro de Ciências Humanas e Naturais, Universidade Federal do Espírito Santo, Maruipe, Vitória, ES 29043-900, Brazil; and dDepartment of Anthropology, University of Texas at Austin, Austin, TX 78712 Contributed by Karen B. Strier, October 12, 2011 (sent for review September 11, 2011) Levels of reproductive skew vary in wild primates living in The fecal samples used for paternity analyses included the 22 multimale groups depending on the degree to which high-ranking infants born between 2005 and 2007 that survived to ≥2.08 y of males monopolize access to females. Still, the factors affecting age, their 21 mothers (representing diverse ages and histories) paternity in egalitarian societies remain unexplored. We combine (Table S1), and the 24 adult males that were possible sires of at unique behavioral, life history, and genetic data to evaluate the least one infant (Table S2). We considered a male to be a po- distribution of paternity in the northern muriqui (Brachyteles tential sire for an infant if he was >5 y and was known to have hypoxanthus), a species known for its affiliative, nonhierarchical completed a copulation before the infant’s estimated conception relationships. We genotyped 67 individuals (22 infants born over a date (birth date minus mean 216.4-d gestation) (16). -

Diets of Howler Monkeys
Chapter 2 Diets of Howler Monkeys Pedro Américo D. Dias and Ariadna Rangel-Negrín Abstract Based on a bibliographical review, we examined the diets of howler mon- keys to compile a comprehensive overview of their food resources and document dietary diversity. Additionally, we analyzed the effects of rainfall, group size, and forest size on dietary variation. Howlers eat nearly all available plant parts in their habitats. Time dedicated to the consumption of different food types varies among species and populations, such that feeding behavior can range from high folivory to high frugivory. Overall, howlers were found to use at least 1,165 plant species, belonging to 479 genera and 111 families as food sources. Similarity in the use of plant taxa as food sources (assessed with the Jaccard index) is higher within than between howler species, although variation in similarity is higher within species. Rainfall patterns, group size, and forest size affect several dimensions of the dietary habits of howlers, such that, for instance, the degree of frugivory increases with increased rainfall and habitat size, but decreases with increasing group size in groups that live in more productive habitats. Moreover, the range of variation in dietary habits correlates positively with variation in rainfall, suggesting that some howler species are habitat generalists and have more variable diets, whereas others are habi- tat specialists and tend to concentrate their diets on certain plant parts. Our results highlight the high degree of dietary fl exibility demonstrated by the genus Alouatta and provide new insights for future research on howler foraging strategies. Resumen Con base en una revisión bibliográfi ca, examinamos las dietas de los monos aulladores para describir exhaustivamente sus recursos alimenticios y la diversidad de su dieta. -

Lippia Adoensis, Implication for Access and Benefit Sharing Agreement in Sidama and Westarsi Zones, SNNPR and Oromia Regions, Ethiopia
MOJ Ecology & Environmental Sciences Research Article Open Access Economic Valuation of Lippia adoensis, implication for Access and Benefit Sharing agreement in Sidama and WestArsi Zones, SNNPR and Oromia Regions, Ethiopia Abstract Volume 4 Issue 5 - 2019 Biodiversity has major economic value. Most of these values are often not captured by the market. Hence, the potential of biodiversity is often underestimated. Such an Amare Seifu, Tesfaye Bekele, Manaye underestimation is considered as one of the factors for rapid depletion of biodiversity Misganawand, Ashenafi Ayenew and loss of habitats and species. Valuation of bio-resources would facilitate in identifying Ethiopian Biodiversity Institute, Genetic Resources Access and the real value of genetic resources and obtaining a reasonably better share of the overall Benefit Sharing Directorate, Ethiopia benefits of genetic materials to the local communities, who are involved in its management. Accordingly, the Economic Valuation of Lippia adoensis was conducted implication for Correspondence: Amare Seifu, Ethiopian Biodiversity Institute, ABS. The result of this study identified the direct and indirect use values ofLippia adoensis. Genetic Resources Access and Benefit Sharing Directorate, Ethiopia, Tel +251916595709, Email Majority of the respondents (98.33%) were willing to pay for use values of Lippia adoensis. Moreover, (80.8%) of the respondents were willingness to pay for Conservation of Lippia Received: October 09, 2019 | Published: October 30, 2019 adoensis. Furthermore, 95(79.2%) of the respondents were willingness to pay for non-use value of Lippia adoensis in the study areas. The cumulative estimates of willingness to pay for use of values Lippia adoensis were 30,437.50 birr with an average annual willingness to pay of 253.64±275.63 birr for Lippia adoensis. -

Controlled Animals
Environment and Sustainable Resource Development Fish and Wildlife Policy Division Controlled Animals Wildlife Regulation, Schedule 5, Part 1-4: Controlled Animals Subject to the Wildlife Act, a person must not be in possession of a wildlife or controlled animal unless authorized by a permit to do so, the animal was lawfully acquired, was lawfully exported from a jurisdiction outside of Alberta and was lawfully imported into Alberta. NOTES: 1 Animals listed in this Schedule, as a general rule, are described in the left hand column by reference to common or descriptive names and in the right hand column by reference to scientific names. But, in the event of any conflict as to the kind of animals that are listed, a scientific name in the right hand column prevails over the corresponding common or descriptive name in the left hand column. 2 Also included in this Schedule is any animal that is the hybrid offspring resulting from the crossing, whether before or after the commencement of this Schedule, of 2 animals at least one of which is or was an animal of a kind that is a controlled animal by virtue of this Schedule. 3 This Schedule excludes all wildlife animals, and therefore if a wildlife animal would, but for this Note, be included in this Schedule, it is hereby excluded from being a controlled animal. Part 1 Mammals (Class Mammalia) 1. AMERICAN OPOSSUMS (Family Didelphidae) Virginia Opossum Didelphis virginiana 2. SHREWS (Family Soricidae) Long-tailed Shrews Genus Sorex Arboreal Brown-toothed Shrew Episoriculus macrurus North American Least Shrew Cryptotis parva Old World Water Shrews Genus Neomys Ussuri White-toothed Shrew Crocidura lasiura Greater White-toothed Shrew Crocidura russula Siberian Shrew Crocidura sibirica Piebald Shrew Diplomesodon pulchellum 3. -

Online Appendix for “The Impact of the “World's 25 Most Endangered
Online appendix for “The impact of the “World’s 25 Most Endangered Primates” list on scientific publications and media” Table A1. List of species included in the Top25 most endangered primate list from the list of 2000-2002 to 2010-2012 and used in the scientific publication analysis. There is the year of their first mention in the Top25 list and the consecutive mentions in the following Top25 lists. Species names are the current species names (based on IUCN) and not the name used at the time of the Top25 list release. First Second Third Fourth Fifth Sixth Species mention mention mention mention mention mention Ateles fusciceps 2006 Ateles hybridus 2006 2008 2010 Ateles hybridus brunneus 2004 Brachyteles hypoxanthus 2000 2002 2004 Callicebus barbarabrownae 2010 Cebus flavius 2010 Cebus xanthosternos 2000 2002 2004 Cercocebus atys lunulatus 2000 2002 2004 Cercocebus galeritus galeritus 2002 Cercocebus sanjei 2000 2002 2004 Cercopithecus roloway 2002 2006 2008 2010 Cercopithecus sclateri 2000 Eulemur cinereiceps 2004 2006 2008 Eulemur flavifrons 2008 2010 Galagoides rondoensis 2006 2008 2010 Gorilla beringei graueri 2010 Gorilla beringei beringei 2000 2002 2004 Gorilla gorilla diehli 2000 2002 2004 2006 2008 Hapalemur aureus 2000 Hapalemur griseus alaotrensis 2000 Hoolock hoolock 2006 2008 Hylobates moloch 2000 Lagothrix flavicauda 2000 2006 2008 2010 Leontopithecus caissara 2000 2002 2004 Leontopithecus chrysopygus 2000 Leontopithecus rosalia 2000 Lepilemur sahamalazensis 2006 Lepilemur septentrionalis 2008 2010 Loris tardigradus nycticeboides -

Ethiopia: West Guji – Gedeo Conflict Displacement Flash Update 2
Ethiopia: West Guji – Gedeo Conflict Displacement Flash Update 2 22 May 2018 Highlights • A rapid assessment conducted in Gedeo zone (SNNPR) found that at least 274,5481 people were displaced from West Guji (Oromia) to Gedeo zone following the violence that erupted along the regional boundary in mid-April. Some 84,000 people were also displaced in West Guji zone. More than 4,000 houses were reportedly burnt down or damaged in West Guji and livelihoods destroyed. • Official reports indicate that as of early May, 102,593 people of the 274,548 people displaced in Gedeo were returned to their villages in West Guji zone • The Government, the local community and civil society were the first and main responders to date, with sup- port from few partners. Significant gaps in food, shelter, NFI and WaSH are reported • Although there were no reported health outbreaks so far, AWD, scabies and measles are potential health risks due to the use of water from unsafe sources, open defecation, overcrowded shelter and sharing of clothes. The overcrowded shelter conditions within hosting households also present protection concerns. Dara Sidama Dila I. Displacement overview ! 12,176 452 Bore Chichu A rapid assessment conducted in Gedeo zone ! 3,772 (SNNPR) from 2 to 8 May found that at least Dila Zuria stite eturns 274,548 people (139,494 male; 135,104 female) Wenago einin ! were displaced from West Guji (Oromia) to Gedeo Bule isceent 4,844 ! Koti ! zone, following the inter-communal violence that 8,198 Ilalcha ! erupted in mid-April 2018 along the borders of the Wenago Bule Abaya two zones.