Reticulocyte Maturation Process — Experimental Demonstration of RET Channel Using Anemic Mice —
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

20 Hemolytic Anemias Due to Abnormal Red Cell Enzymes
Hemolytic Anemias Due to Abnormal Red Cell Enzymes MODULE Hematology and Blood Bank Technique 20 HEMOLYTIC ANEMIAS DUE TO Notes ABNORMAL RED CELL ENZYMES 20.1 INTRODUCTION The main metabolic substrate for the RBCs is glucose. It is metabolized by two pathways: approximately 90% of the glucose is metabolized through the Embden Meyerhoff (glycolytic) pathway and the rest by the hexose monophosphate (HMP) pathway. In the Embden Meyerhoff (glycolytic) pathway glucose is metabolized to lactate through a series of enzymatic steps. Each molecule of glucose gives rise to 2 molecules of ATP. The ATP provides energy to maintain red cell volume, shape and flexibility. An ATP dependent pump in the red cell membrane actively keeps sodium out of the cell and potassium inside. The red cell has the enzymes that are needed for the glycolytic pathway. These enzymes help break down glucose to generate ATP which is the source of energy. About 10% of the glucose is diverted to the Hexose Monophosphate shunt pathway and this is essential for protection of red cells from oxidative stress. This pathway is necessary for the generation of NADPH which then reduces oxidized glutathione (GSSG) to reduced glutathione (GSH). GSH prevents the accumulation of H2O2 and the oxidation of hemoglobin to methemoglobin. When the level of GSH falls, H2O2 accumulates in the cell and oxidizes the hemoglobin to methemoglobin which becomes denatured and precipitates as Heinz bodies. These inclusions are rigid and attached to the red cell membrane and make the red cell susceptible to hemolysis. The NADPH required in this pathway is generated by the enzyme Glucose 6 phosphate dehydrogenase (G6PD). -
Hematology Unit Lab 1 Review Material
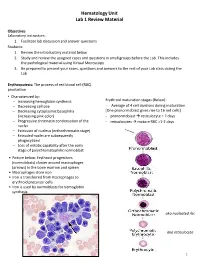
Hematology Unit Lab 1 Review Material Objectives Laboratory instructors: 1. Facilitate lab discussion and answer questions Students: 1. Review the introductory material below 2. Study and review the assigned cases and questions in small groups before the Lab. This includes the pathological material using Virtual Microscopy 3. Be prepared to present your cases, questions and answers to the rest of your Lab class during the Lab Erythropoiesis: The process of red blood cell (RBC) production • Characterized by: − Increasing hemoglobin synthesis Erythroid maturation stages (Below): − Decreasing cell size - Average of 4 cell divisions during maturation − Decreasing cytoplasmic basophilia [One pronormoblast gives rise to 16 red cells] (increasing pink color) - pronormoblast → reticulocyte = 7 days − Progressive chromatin condensation of the - reticulocytes → mature RBC =1-2 days nuclei − Extrusion of nucleus (orthochromatic stage) − Extruded nuclei are subsequently phagocytized − Loss of mitotic capability after the early stage of polychromatophilic normoblast • Picture below: Erythroid progenitors (normoblasts) cluster around macrophages (arrows) in the bone marrow and spleen • Macrophages store iron • Iron is transferred from macrophages to erythroid precursor cells • Iron is used by normoblasts for hemoglobin synthesis aka nucleated rbc aka reticulocyte 1 Mature Red Blood Cell 7-8 microns; round / ovoid biconcave disc with orange-red cytoplasm, no RNA, no nucleus; survives ~120 days in circulation Classification of Anemia by Morphology 1. -

Basic Hematology Overview
1 BASIC HEMATOLOGY OVERVIEW Name___________________________ 1. Mean cell volume (MCV) is calculated using the following formulas: a) hgb/rbc x 10 b) hct/rbc x 10 c) hct/hgb x 100 d) hgb/rbc x 100 2. Calculate the mean cell hemoglobin concentration (MCHC) using the following values: Hgb: 15 g/dl (150g/L); RBC: 4.50 x 10^6/ul (4.50 x 10^12/L) Hct: 47% (0.47) a) 9.5% (0.95) b) 10.4% (.104) c) 31.9% (.319) d) 33.3% (.333) 3. A manual white blood cell (WBC) count was performed. A total of 36 cells were counted in all 9 mm^2 squares of a Neubauer-ruled hematocytometer. A dilution 1:10 dilution was used. What is the Wbc count? a) 0.4 x 10^9/L b) 2.5 x 10^9/L c) 4.0 x 10^9/L D) 8.0 x 10^9/L Prepared by Julia Fonseca Melendez, BS, MT (ASCP), SH(ACP), SCYM(ASCP) 2 BASIC HEMATOLOGY OVERVIEW Name__________________________ 4. A 7.0 ml etheylenediaminetetraacetic acid (EDTA) tube is received in the laboratory containing only 2.0 ml of blood. If the laboratory is using manual techniques, which of the following tests will most likely be erroneous? a) RBC count b) Hgb c) Hematocrit d) Wbc count 5. A decreased osmotic fragility test would be associated with which of the following conditions? a) Sickle cell anemia b) Hereditary spherocytosis c) Hemolytic disease of the newborn d) Acquired hemolytic anemia 6. A falsely elevated hematocrit is obtained using a defective centrifuge. -

BRILLIANT CRESYL BLUE Powder Dye, C.I. 51010
BRILLIANT CRESYL BLUE powder dye, C.I. 51010 IVD In vitro diagnostic medical device Brilliant Blue C For use in hematology and histology INSTRUCTIONS FOR USE REF Product code: BCB-P-10 (10 g) BCB-P-25 (25 g) Introduction Supravital staining is a method of staining used in microscopy to examine living cells that have been removed from an organism. It differs from intravital staining, which is done by injecting or otherwise introducing the stain into the body. Thus a supravital stain may have a greater toxicity, as only a few cells need to survive it for a short period of time. As the cells are alive and unfixed, outside the body, supravital stains are temporary in nature. The most common supravital stain is performed on reticulocytes using brilliant cresyl blue, which makes it possible to see the reticulofilamentous pattern of ribosomes characteristically precipitated in these live immature red blood cells by the supravital stains. By counting the number of such cells the rate of red blood cell formation can be determined, providing an insight into bone marrow activity and anemia. Product description BRILLIANT CRESYL BLUE - Powder dye for manufacturing solution for microscopic counting of reticulocytes and for other uses in histology Other preparations and reagents used in preparing the dye solution: Saline (0.85% NaCl) Preparing the solutions for staining Brilliant Cresyl Blue, 1% solution: Dilute 1 g of Brilliant Cresyl Blue powder dye in 100 mL of saline and filter Store as saturated (stock) solution Prepare the working solution in form of diluted stock dye solution and saline in ratios ranging from 1:80 to 1:200. -

The Color of Tissue Diagnostics Routine Stains, Special Stains and Ancillary Reagents
The Color of Tissue Diagnostics Routine Stains, Special Stains and Ancillary Reagents The life science business of Merck KGaA, Darmstadt, Germany operates as MilliporeSigma in the U.S. and Canada. For over years, 100routine stains, special stains and ancillary reagents have been part of the MilliporeSigma product range. This tradition and experience has made MilliporeSigma one of the world’s leading suppliers of microscopy products. The products for microscopy, a comprehensive range for classical hematology, histology, cytology, and microbiology, are constantly being expanded and adapted to the needs of the user and to comply with all relevant global regulations. Many of MilliporeSigma’s microscopy products are classified as in vitro diagnostic (IVD) medical devices. Quality Means Trust As a result of MilliporeSigma’s focus on quality control, microscopy products are renowned for excellent reproducibility of results. MilliporeSigma products are manufactured in accordance with a quality management system using raw materials and solvents that meet the most stringent quality criteria. Prior to releasing the products for particular applications, relevant chemical and physical parameters are checked along with product functionality. The methods used for testing comply with international standards. For over Contents Ancillary Reagents Microbiology 3-4 Fixing Media 28-29 Staining Solutions and Kits years, 5-6 Embedding Media 30 Staining of Mycobacteria 100 6 Decalcifiers and Tissue Softeners 30 Control Slides 7 Mounting Media Cytology 8 Immersion -

E-Learn LAB — RCD 1708
e-Learn LAB — Hemoglobinopathy Based on IQMH Centre for Proficiency Testing Survey RCD-1708-WB Confidence. Elevated. © Institute for Quality Management in Healthcare 1 Disclaimer and Copyright Disclaimer This document contains content developed by IQMH. IQMH’s work is guided by the current best available evidence at the time of publication. The application and use of this document is the responsibility of the user, and IQMH assumes no liability resulting from any such application or use. This document may be reproduced without permission for non-commercial purposes only and provided that appropriate credit is given to IQMH. Copyright The reader is cautioned not to take any single item, or part thereof, of this document out of context. Information presented in this document is the sole property and copyright of the Institute for Quality Management in Healthcare (IQMH). The logos and/or symbols used are the property of IQMH or other third parties. © Institute for Quality Management in Healthcare. All rights reserved. Confidence. Elevated. © Institute for Quality Management in Healthcare 2 Focus of this Presentation • This is a case study of a hemoglobinopathy investigation to familiarize participants with the laboratory features of the unstable hemoglobins, as well as the pathophysiology of common red cell inclusions. • You will be presented with clinical information, images, and laboratory results and will be prompted with self- learning questions. Confidence. Elevated. © Institute for Quality Management in Healthcare 3 Credits Case and discussion provided by members of the 2017 IQMH Hematology Scientific Committee, and the IQMH Consultant Technologist. Confidence. Elevated. © Institute for Quality Management in Healthcare 4 Clinical Information A 14-year-old female is being assessed for gall stones. -
A Laboratory Guide to Clinical Hematology
A Laboratory Guide to Clinical Hematology A Laboratory Guide to Clinical Hematology A Laboratory Guide to Clinical Hematology VALENTIN VILLATORO AND MICHELLE TO EDMONTON A Laboratory Guide to Clinical Hematology by Michelle To is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted. Please be aware that the content for the entirety of this eBook is subject to a creative common license: Attribution-NonCommercial 4.0 International (CC BY-NC 4.0) You are free to: Share — copy and redistribute the material in any medium or format Adapt — remix, transform, and build upon the material The licensor cannot revoke these freedoms as long as you follow the license terms. Under the following terms: Attribution — You must give appropriate credit, provide a link to the license, and indicate if changes were made. You may do so in any reasonable manner, but not in any way that suggests the licensor endorses you or your use. NonCommercial — You may not use the material for commercial purposes. No additional restrictions — You may not apply legal terms or technological measures that legally restrict others from doing anything the license permits. Contents Authors & Editors ................................................................................................................................... xii Creative Commons License and Citation ............................................................................................... xiii Contact Information and Feedback ........................................................................................................ -

Heinz Body Anaemia in Two Dogs After Catalan Spring Onion (“Calcot”) Ingestion: a Case Reports
Case Report Veterinarni Medicina, 53, 2008 (7): 392–395 Heinz body anaemia in two dogs after Catalan spring onion (“calcot”) ingestion: a case reports R. Guitart1, C. Mateu2, A. Lopez i Agullo3, J. Alberola1 1Autonomous University of Barcelona, Bellaterra, Spain 2Veterinary Hospital of the Maresme, Mataro, Spain 3Veterinary Services of Mataro, Mataro, Spain ABSTRACT: Catalan spring onions, or “calcots”, are a very popular kind of vegetables obtained by special agro- nomic practices that avoid photosynthesis and subsequent metabolism by the Allium cepa plants. They have been considered for more than a century harmless for pets, as pleasant odour and sweet taste differ so largely from any other onion that can be ingested by children and adults in huge quantities, up to ~3% body weight, without problems being observed. However, the organosulphur chemicals responsible for oxidative damage to canine red blood cells found in this plant species seem to remain present, as two dogs were confirmed to be poisoned after eating left-over “calcots”. Both patients presented haemolytic anaemia with Heinz body formation, and eccent- rocytosis. Veterinarians and owners should be aware of the dangers associated with this particular onion variety and type, as it is highly palatable to pets and no effective treatment is currently available. Keywords: Allium cepa; poisoning; organosulphur compounds; oxidative damage The onion (Allium cepa) is one of the oldest Catalan spring onion, locally and internationally crops. Native to Central Asia (Brown, 2002), on- known as “calcot”, is a variety of A. cepa known as ion and garlic (A. sativum) are probably the most Blanca Gran Tardana cultivated mainly in Catalonia well-known representatives of the genus and also (North East of Spain), under very strict conditions.