Attraction of Nocturnally Migrating Birds to Artificial Light the Influence
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

345 Fieldfare Put Your Logo Here
Javier Blasco-Zumeta & Gerd-Michael Heinze Sponsor is needed. Write your name here Put your logo here 345 Fieldfare Fieldfare. Adult. Male (09-I). Song Thrush FIELDFARE (Turdus pilaris ) IDENTIFICATION 25-26 cm. Grey head; red-brown back; grey rump and dark tail; pale underparts; pale flanks spotted black; white underwing coverts; yellow bill with ochre tip. Redwing Fieldfare. Pattern of head, underwing co- verts and flank. SIMILAR SPECIES Song Thrush has orange underwing coverts; Redwing has reddish underwing coverts; Mistle Thrush has white underwing coverts, but lacks pale supercilium and its rump isn’t grey. Mistle Thrush http://blascozumeta.com Write your website here Page 1 Javier Blasco-Zumeta & Gerd-Michael Heinze Sponsor is needed. Write your name here Put your logo here 345 Fieldfare SEXING Male with dark or black tail feathers; red- dish feathers on back with blackish center; most have a broad mark on crown feathers. Female with dark brown tail feathers but not black; dull reddish feathers on back with dark centre (but not blackish); most have a thin mark on crown feathers. CAUTION: some birds of both sexes have similar pattern on crown feathers. Fieldfare. Sexing. Pattern of tail: left male; right fe- male. Fieldfare. Sexing. Pat- AGEING tern of Since this species doesn’t breed in Aragon, only crown feat- 2 age groups can be recognized: hers: top 1st year autumn/2nd year spring with moult male; bot- limit within moulted chestnut inner greater co- tom female. verts and retained juvenile outer greater coverts, shorter and duller with traces of white tips; pointed tail feathers. -

New Zealand Comprehensive II Trip Report 31St October to 16Th November 2016 (17 Days)
New Zealand Comprehensive II Trip Report 31st October to 16th November 2016 (17 days) The Critically Endangered South Island Takahe by Erik Forsyth Trip report compiled by Tour Leader: Erik Forsyth RBL New Zealand – Comprehensive II Trip Report 2016 2 Tour Summary New Zealand is a must for the serious seabird enthusiast. Not only will you see a variety of albatross, petrels and shearwaters, there are multiple- chances of getting out on the high seas and finding something unusual. Seabirds dominate this tour and views of most birds are alongside the boat. There are also several land birds which are unique to these islands: kiwis - terrestrial nocturnal inhabitants, the huge swamp hen-like Takahe - prehistoric in its looks and movements, and wattlebirds, the saddlebacks and Kokako - poor flyers with short wings Salvin’s Albatross by Erik Forsyth which bound along the branches and on the ground. On this tour we had so many highlights, including close encounters with North Island, South Island and Little Spotted Kiwi, Wandering, Northern and Southern Royal, Black-browed, Shy, Salvin’s and Chatham Albatrosses, Mottled and Black Petrels, Buller’s and Hutton’s Shearwater and South Island Takahe, North Island Kokako, the tiny Rifleman and the very cute New Zealand (South Island wren) Rockwren. With a few members of the group already at the hotel (the afternoon before the tour started), we jumped into our van and drove to the nearby Puketutu Island. Here we had a good introduction to New Zealand birding. Arriving at a bay, the canals were teeming with Black Swans, Australasian Shovelers, Mallard and several White-faced Herons. -

0854 BC Annual Review 04
ann ual review for 2010/11 Saving butte rflies, moths and our environment Highlights of the year Overview by Chairman and Chief Executive In this Annual Review, we celebrate our achievements over the last year and look Several of our most threatened butterflies and moths Secured a core funding grant for Butterfly Conservation ahead to explain our ambitious “2020 vision” for the current decade and beyond. began to recover thanks to our landscape scale projects. Europe from the EU, which enabled the employment of staff for the first time. Successes include the Pearl- bordered Fritillary, Undoubtedly, the most significant success during 2010 is that several of our During the year, we have successfully concluded two of our biggest ever High Brown Fritillary, Duke of Burgundy, Wood White, most threatened species showed signs of recovery directly due to management projects (Moths Count and the South-East Woodlands project) and taken out Small Blue, Grey Carpet and Forester moths. Raised funds to continue our work to save threatened carried out as part of our landscape scale initiatives. Against the background leases on three important new reserves. Our success has been demonstrated species in Scotland, Wales and N. Ireland and for major of decades of decline and habitat loss, these increases show that our by a growth in membership to almost 16,000 and by the continuing support Acquired three new reserves which support important new landscape projects to save the Duke of Burgundy conservation strategy is working. of those members who have responded generously to our appeals and populations of threatened species: Myers Allotment on the South Downs and the Large Blue in Somerset. -

Developing Methods for the Field Survey and Monitoring of Breeding Short-Eared Owls (Asio Flammeus) in the UK: Final Report from Pilot Fieldwork in 2006 and 2007
BTO Research Report No. 496 Developing methods for the field survey and monitoring of breeding Short-eared owls (Asio flammeus) in the UK: Final report from pilot fieldwork in 2006 and 2007 A report to Scottish Natural Heritage Ref: 14652 Authors John Calladine, Graeme Garner and Chris Wernham February 2008 BTO Scotland School of Biological and Environmental Sciences, University of Stirling, Stirling, FK9 4LA Registered Charity No. SC039193 ii CONTENTS LIST OF TABLES................................................................................................................... iii LIST OF FIGURES ...................................................................................................................v LIST OF FIGURES ...................................................................................................................v LIST OF APPENDICES...........................................................................................................vi SUMMARY.............................................................................................................................vii EXECUTIVE SUMMARY ................................................................................................... viii CRYNODEB............................................................................................................................xii ACKNOWLEDGEMENTS....................................................................................................xvi 1. BACKGROUND AND AIMS...........................................................................................2 -

Biodiversity Information Report 13/07/2018
Biodiversity Information Report 13/07/2018 MBB reference: 2614-ARUP Site: Land near Hermitage Green Merseyside BioBank, The Local Biodiversity Estate Barn, Court Hey Park Roby Road, Liverpool Records Centre L16 3NA for North Merseyside Tel: 0151 737 4150 [email protected] Your Ref: None supplied MBB Ref: 2614-ARUP Date: 13/07/2018 Your contact: Amy Martin MBB Contact: Ben Deed Merseyside BioBank biodiversity information report These are the results of your data request relating to an area at Land near Hermitage Green defined by a buffer of 2000 metres around a site described by a boundary you supplied to us (at SJ598944). You have been supplied with the following: records of protected taxa that intersect the search area records of BAP taxa that intersect the search area records of Red Listed taxa that intersect the search area records of other ‘notable’ taxa that intersect the search area records of WCA schedule 9 taxa (including ‘invasive plants’) that intersect the search area a map showing the location of monad and tetrad references that overlap the search area a list of all designated sites that intersect your search area citations, where available, for intersecting Local Wildlife Sites a list of other sites of interest (e.g. Ancient Woodlands) that intersect your search area a map showing such sites a list of all BAP habitats which intersect the search area a map showing BAP habitats a summary of the area for all available mapped Phase 1 and/or NVC habitats found within 500m of your site a map showing such habitats Merseyside BioBank (MBB) is the Local Environmental Records Centre (LERC) for North Merseyside. -

EUROPEAN BIRDS of CONSERVATION CONCERN Populations, Trends and National Responsibilities
EUROPEAN BIRDS OF CONSERVATION CONCERN Populations, trends and national responsibilities COMPILED BY ANNA STANEVA AND IAN BURFIELD WITH SPONSORSHIP FROM CONTENTS Introduction 4 86 ITALY References 9 89 KOSOVO ALBANIA 10 92 LATVIA ANDORRA 14 95 LIECHTENSTEIN ARMENIA 16 97 LITHUANIA AUSTRIA 19 100 LUXEMBOURG AZERBAIJAN 22 102 MACEDONIA BELARUS 26 105 MALTA BELGIUM 29 107 MOLDOVA BOSNIA AND HERZEGOVINA 32 110 MONTENEGRO BULGARIA 35 113 NETHERLANDS CROATIA 39 116 NORWAY CYPRUS 42 119 POLAND CZECH REPUBLIC 45 122 PORTUGAL DENMARK 48 125 ROMANIA ESTONIA 51 128 RUSSIA BirdLife Europe and Central Asia is a partnership of 48 national conservation organisations and a leader in bird conservation. Our unique local to global FAROE ISLANDS DENMARK 54 132 SERBIA approach enables us to deliver high impact and long term conservation for the beneit of nature and people. BirdLife Europe and Central Asia is one of FINLAND 56 135 SLOVAKIA the six regional secretariats that compose BirdLife International. Based in Brus- sels, it supports the European and Central Asian Partnership and is present FRANCE 60 138 SLOVENIA in 47 countries including all EU Member States. With more than 4,100 staf in Europe, two million members and tens of thousands of skilled volunteers, GEORGIA 64 141 SPAIN BirdLife Europe and Central Asia, together with its national partners, owns or manages more than 6,000 nature sites totaling 320,000 hectares. GERMANY 67 145 SWEDEN GIBRALTAR UNITED KINGDOM 71 148 SWITZERLAND GREECE 72 151 TURKEY GREENLAND DENMARK 76 155 UKRAINE HUNGARY 78 159 UNITED KINGDOM ICELAND 81 162 European population sizes and trends STICHTING BIRDLIFE EUROPE GRATEFULLY ACKNOWLEDGES FINANCIAL SUPPORT FROM THE EUROPEAN COMMISSION. -

Stratford Park Bird Report 2019 Mike Mccrea (Contract Supervisor)
Stratford Park Bird Report 2019 Mike McCrea (Contract Supervisor) (Common Buzzard in Stratford Park Photo: Mike McCrea) Hi All, This month’s Biodiversity Newsletter features a Stratford Park Bird Report for 2019. Mike INTRODUCTION Birds globally are facing increasing challenges; the main drivers being Climate change, agricultural management, hydrological change, urbanisation and invasive non-native species. These factors in the last thirty years have caused the world’s bird populations to decline by 40%. Birds are an indicator species – their presence (or lack thereof) is a sign of the good health (or otherwise) of an ecosystem. The recently published Wild Bird Populations in the UK 1970 – 2018 report shows that many of our familiar birds are still declining, with several doing so at a worrying rate. This downward trend is happening globally in a wide range of habitats. Within the context of global bird distribution, Stratford Park is but a microcosm within the whole picture but as important as any other habitat for wild birds, for it is this park, like many others throughout the UK, which provides important habitats for our birds. Within a snapshot of 30 years the park has lost at least six species of birds, several of which were previously common – bullfinch and spotted flycatcher being the most notable. Others such as Mistle thrush and song thrush have declined at an alarming rate. During the last 10 years we have helped the recovery of many of the park’s bird species through the introduction of next boxes and creating new habitats. The Stratford Park Biodiversity & Landscape Action Plan (2011) has been the catalyst for biodiversity development and a working tool which we have used to identify key areas throughout the park that require conservation measures for the benefit of birds and other animals. -

Birds: Structure, Function and Adaptation
Birds: Structure, function and adaptation Birds: Structure, function and adaptation This interactive diagram explores the sequential and interlinking science concepts that underpin knowledge and understanding about birds’ physical features, their functions and how they help birds with flight, feeding and life in particular habitats. The concepts listed just above the overarching concepts reflect learning at New Zealand Curriculum level 1 and show how they may build in sequence to level 4. The overarching science concepts are fully developed concepts and might not be achieved until level 7 or 8. Some of the text is courtesy of the New Zealand Ministry of Education’s Building Science Concepts Book 3 Birds: Structure, Function, and Adaptation. The links to Hub resources provide additional background information and classroom activities that will support teachers to scaffold the development of their students’ conceptual understanding about birds. The images provide a means to initiate discussions, check student thinking and consolidate student understanding. The article Building Science Concepts: Birds provides additional science and pedagogical information. Index • Highly specialised birds have unique adaptations that restrict them to habitats that meet their needs • Birds that are common in towns have adaptations that enable them to cope with changes brought about by people • We can usually tell what sort of food a bird eats by looking at its beak and feet • All birds have feathers, two legs and a beak instead of teeth © Copyright. Science -

Adobe PDF, Job 6
Noms français des oiseaux du Monde par la Commission internationale des noms français des oiseaux (CINFO) composée de Pierre DEVILLERS, Henri OUELLET, Édouard BENITO-ESPINAL, Roseline BEUDELS, Roger CRUON, Normand DAVID, Christian ÉRARD, Michel GOSSELIN, Gilles SEUTIN Éd. MultiMondes Inc., Sainte-Foy, Québec & Éd. Chabaud, Bayonne, France, 1993, 1re éd. ISBN 2-87749035-1 & avec le concours de Stéphane POPINET pour les noms anglais, d'après Distribution and Taxonomy of Birds of the World par C. G. SIBLEY & B. L. MONROE Yale University Press, New Haven and London, 1990 ISBN 2-87749035-1 Source : http://perso.club-internet.fr/alfosse/cinfo.htm Nouvelle adresse : http://listoiseauxmonde.multimania. -

Breeding Biology of Icelandic Thrushes
Breeding biology of Icelandic thrushes Hulda Elísabet Harðardóttir Faculty of Life and Environmental Sciences University of Iceland 2019 Breeding biology of Icelandic thrushes Hulda Elísabet Harðardóttir 90 ECTS thesis submitted in partial fulfillment of a Magister Scientiarum degree in Biology MS Committee Gunnar Þór Hallgrímsson Snæbjörn Pálsson Master’s Examiner Tómas Grétar Gunnarsson Faculty of Life and Environmental Sciences School of Engineering and Natural Sciences University of Iceland Reykjavik, May 2019 Breeding biology of Icelandic thrushes Biology of thrushes 90 ECTS thesis submitted in partial fulfillment of a Magister Scientiarum degree in Biology Copyright © 2019 Hulda Elísabet Harðardóttir All rights reserved Faculty of Life and Environmental Sciences School of Engineering and Natural Sciences University of Iceland Askja, Sturlugata 7 107, Reykjavik Iceland Telephone: 525 4000 Bibliographic information: Hulda Elísabet Harðardóttir, 2019, Breeding biology of Icelandic thrushes, Master’s thesis, Faculty of Life and Environmental Sciences, University of Iceland, pp. 68. Printing: Háskólaprent Reykjavik, Iceland, May 2019 Abstract This thesis integrates in three separate chapters some aspects of the breeding biology of the Eurasian redwing (Turdus iliacus coburni) and the Eurasian blackbird (T. merula). The redwing has been breeding in Iceland for centuries while the blackbird colonized between 1990-2000 and is still a relatively scarce breeding bird outside SW-Iceland. Both species are understudied in Iceland despite many valuable research questions, especially on interactions between the species due to the recent colonisation of the blackbird. The study was conducted in two consecutive summers in 2017 and 2018, in Fossvogur cemetery located centrally in Reykjavik Iceland. The first chapter focuses on various aspects of their breeding biology including timing, nest site selection and breeding success. -

Common Urban Birds
Common Urban Birds Crested Pigeon Spotted Turtle Dove* Feral Pigeon* Noisy Miner New Holland Eastern Spinebill White-plumed Honeyeater Honeyeater JS SW SW SW SW JS JS (Crest on head) (White spots on neck) (Dark grey feathers usually with a (Black head, yellow around eyes) (Black and yellow wings, Black and (Black, white and reddish-brown (White lines on neck) shiny green neck) white striped chest) feathers) Nectarivore & Granivore Granivore Granivore Nectarivore & Insectivore Nectarivore & Insectivore Nectarivore & Insectivore q q q q Insectivore,Omnivore q q q X Ground X Trees,Shrubs,Ground X Ground X Trees,Shrubs,Ground,Air X Trees,Shrubs,Air X Shrubs,Air X Trees,Shrubs,Ground,Air Red Wattlebird Little Wattlebird Striated Pardalote Welcome Swallow House Sparrow* Silvereye Willie Wagtail JS SW JH SW JMG JT JS (Yellow-orange belly, red wattles) (No orange on belly, no wattles) (Yellow face, black & white (Flies around ovals and other (Very small) (Silver ring around eye) (Black and white, tail wags from streaked crown, white wing streaks grassed areas, forked tail) side to side) with red spot) Nectarivore & Nectarivore & Insectivore Nectarivore & Insectivore Insectivore Granivore Omnivore Insectivore q q q Insectivore,Insectivore q q q q X Trees,Shrubs,Air X Trees,Shrubs,Air X Trees,Shrubs X Air X Ground X Trees,Shrubs X Ground,Air Common Blackbird* Common Starling* Australian Magpie Magpie-lark Little Raven Laughing Nankeen Kestrel Kookaburra JS JS JG JS JS JG JS (Smaller beak and body than (Breeding male black with bright yellow (Dark -
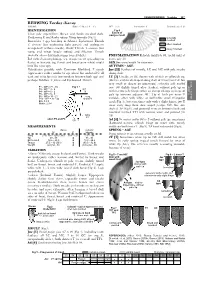
REDWING Turdus Iliacus (REDWI) Ring: 3.5 MA (3.8 – 4.2) WP = 3 (4) Incubation: F Parental Care: F, M IDENTIFICATION Fig 4 Extent of Clear Pale Supercilium
PASSERIFORMES – Turdidae 267 REDWING Turdus iliacus (REDWI) Ring: 3.5 MA (3.8 – 4.2) WP = 3 (4) Incubation: F Parental care: F, M IDENTIFICATION Fig 4 Extent of Clear pale supercilium. Breast and flanks streaked dark. postjuv moult Underwing C and flanks rufous. Wing formula (Fig 1). generally Resembles 3 spp breeding in Siberia: Eyebrowed Thrush moulted T. obscurus (but underwing light greyish and underparts often moulted orange-buff without streaks), Dusky Thrush T. e u n om u s (but rarely moulted rump and wings largely rufous) and Siberian Thrush Geokichla sibirica (but underwings barred white). PNEUMATISATION Reliable until late 09, useful until at Ind with aberrant plumage (eg, orange breast spreading on least early 10. flanks) or leucistic (eg, throat and breast pure white) might SEX See wing length for extremes. look like rarer spp. AUTUMN – AGE Hybridisation possible with Common Blackbird T. m e r u la Juv [3J] Feathers of mantle, LC and MC with pale streaks (appearance rather similar to ssp coburni but undertail C all along shaft. dark and wing formula intermediate between both spp) and 1Y [3] Usually, juv GC shorter with whitish or yellowish tip, perhaps Fieldfare T. pilaris and Eyebrowed Thrush. (distinct and streak shaped along shaft of innermost GC but very small or absent on outermost) contrasting with moulted P1 - WP = 71 - 85 Fig 1 inner GC slightly tinged olive (darker), without pale tip or TURVIS P2 - WP = 3 - 8 ≤ 8 - – P4 - WP = 0 - 2 16 with narrow pale fringe: often an abrupt change in shape of P5 - WP = 5 - 8 pale tip between adjacent GC.