Prosthechea: a Chemical Discontinuity in Laeliinae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Genus Brassavola, (L.) R.Br
The Genus Brassavola, (L.) R.Br. in W.T.Aiton, Hortus Kew. 5: 216 (1813) Type: Brassavola [B.] cucullata [bra-SAH-vo-la kyoo-kyoo-LAH-ta] There are 28 species (OrchidWiz [update Dec 2017]) that are epiphytes and sometimes lithophytes at elevations of from sea level to 3300 ft (1000 m) from Mexico, southern Caribbean islands to northern Argentina in moist or wet montane forests, mangroves, rocky crevices and cliff faces. They are most fragrant at night and many with a citrus smell. The genus is characterized by very small pencil-like pseudobulbs, often forming large clumps; a single, fleshy, apical, sub-terete leaf and the inflorescence produced form the apex of the pseudobulb. The inflorescence carries from a single to a few large flowers. The floral characteristics are elongate narrow similar sepals and petals, the base of the lip usually tightly rolled around at least a portion of the column which carries 12, sometimes eight unequal pollina with prominent opaque caudicles. The flowers usually occur, as a rule, in spring, summer and fall. The flowers are generally yellow to greenish white with a mostly white lip. It is not unusual for dark spots, usually purple, to be in the region where the sepals, petals, and lip join the stem (claw). This spotting is a dominant generic trait in Brassavola nodose. They are easily cultivated under intermediate conditions. Although this is a relatively small genus (28 species), the species show an unusually close relationship with one another in their floral patterns, coloration, and column structure making identification difficult, key to know where the plants were collected. -

Epicladium Small Or Guarianthe Dressler & We Higgins (Orchidaceae)?
LANKESTERIANA 5(1): 63-67. 2005. EPICLADIUM SMALL OR GUARIANTHE DRESSLER & W.E. HIGGINS (ORCHIDACEAE)? JOHN BECKNER Curator Orchid Identification Center, Marie Selby Botanical Gardens, 811 South Palm Ave., Sarasota, Florida 32667, USA. [email protected] ABSTRACT. Guarianthe Dressler & W.E.Higgins as a genus has found little favor with orchid growers. If these species are a distinct genus from Cattleya Lindl., under any name, the effects will be substantial. All taxa of this alliance are of considerable horticultural importance, as species and in the ancestry of thousands of garden hybrids. In any case, Guarianthe cannot be replaced by Epicladium Small, which was based on Epidendrum boothianum. RESUMEN. El nombre genérico Guarianthe Dressler & W.E.Higgins ha sido poco aceptado por los horticultores. Si se trata de un género diferente a Cattleya Lindl., este hecho es significativo. Todas las especies de Guarianthe tienen gran importancia hortícola y han dado origen a miles de híbridos artificiales. En todo caso, Guarianthe no puede ser reemplazado por Epicladium Small, el cual se basa en Epidendrum boothianum. Ambos nombres genéricos no son equivalentes. KEY WORDS / PALABRAS CLAVE: Cattleya, Epicladium, Guarianthe, botanical nomenclature. Julian Shaw, Royal Horticulture Society Senior names, merely numbers, letters, and brief diagnosis. Registrar for Orchid Hybrids, has announced (Shaw They are groups of species that are rather arbitrary, 2004) that: “Guarianthe Predated by Epicladium. and there are no types singled out, so they need not Epicladium (Lindl.) Small is the name with date influence later nomenclature. Of the species Lindley priority for the generic concept recently described gives under Epidendrum, only E. -

Prosthechea Green Hornet
ORCHIDS WE SHOULD ALL GROW by Roy Kreugor PROSTHECHEA GREEN HORNET This particular little gem is a primary hybrid of two well known and loved “cockle shell” orchids, cochlea and lanceifolia. The family tree is a little more complicated with name changes in mid stream. First members of the Epidendrum family, this group was moved to the Encyclia heading when epis and encyclias were split. Later, the separate genus Prosthechea was created for the “cockle shell” type orchids. Today, there are about 100 species in this group, found mostly form Florida to Brazil. The plants are sympodial and mostly epiphytes. The distinction all carry is the shell shaped lip, often striped like a cockle shell, and always found at the top of the flower. Green Hornet is a primary hybrid (first generation cross of two species) of Psh cochlea X Psh lancefolia. It is an easy plant to grow in cultivation, easier than either parent. It takes its shape and size from cochlea, and its color, green form lancefolia. Plants can grow to 24” when mature with a typical size of 12-18”. Flowering season is spring to fall, although some will flower almost continuously. Flowers are borne on slender inflorescences up to 18”tall with 4-12 individual flowers opening almost simultaneously. Flowers are typically 3-4”long with narrow pedals and sepals and a shell shaped striped lip. The lip is a dark maroon on the outer side with a cream inner side striped with fine maroon markings. Pedals and sepals are a light green. Each bloom lasts from several weeks to a month. -

Constituents of Encyclia Longifolia Schltr.(Orchidaceae)
Revista Brasileira de Farmacognosia Brazilian Journal of Pharmacognosy 16(1): 22-23, Jan./Mar. 2006 Received 12/30/05. Accepted 02/17/06 Constituents of Encyclia longifolia Schltr. (Orchidaceae) Jnanabrata Bhattacharyya1*, Abdumalik A. Nishonov1, Leonardo Pessoa Felix2, 3 1 Artigo Maria F.O. Pires , George M. Majetich 1Department of Chemistry, University of Georgia, Athens, GA 30605, USA, 2Departamento de Fitociencia, UFPB, 58397-000, Areia, PB, Brazil, 3Centro de Ciências Biológicas e Agrárias, Universidade Estadual do Piauí, Rua João Cabral s/n, Caixa-Postal 381, Pirajá, 64002-150, Teresina, PI, Brazil RESUMO: “Constituintes de Encyclia longifolia Schltr. (Orchidaceae)”. Uma análise química detalhada dos metabólitos secundários não voláteis de Encyclia longifolia Schltr. Sin. Epidendrum longifolium Barb. Rodr. foi realizada na tentativa de prestar suporte quimiotaxonômico para a recente separação do gênero Encyclia em Encyclia Hook e Prosthechea Knowles & Westc. Unitermos: Encyclia longifolia, Orchidaceae, Prosthechea, 9,10-diidrofenantreno. ABSTRACT: A detailed chemical analysis of the non-volatile secondary metabolites of Encyclia longifolia Schltr. syn. Epidendrum longifolium Barb. Rodr. was carried out in an attempt to lend a chemotaxonomic support for the recent separation of the genus Encyclia into Encyclia Hook and Prosthechea Knowles & Westc. Keywords: Encyclia longifolia, Orchidaceae, Prosthechea, 9, 10-dihydrophenanthrene. INTRODUCTION to isolate and characterize the non-volatile secondary metabolites of several species of -

Epidendrum Secundum (Orchidaceae)
Plant Biology ISSN 1435-8603 RESEARCH PAPER Reproductive biology and pollination mechanisms of Epidendrum secundum (Orchidaceae). Floral variation: a consequence of natural hybridization? E. R. Pansarin & M. C. E. Amaral Departamento de Botaˆ nica, Instituto de Biologia, Universidade Estadual de Campinas, Sa˜ o Paulo, Brazil Keywords ABSTRACT Epidendroideae; Epidendrum; Laeliinae; Orchidaceae; pollination; reproductive biology. The phenology, flower morphology, pollination mechanism and reproductive biology of Epidendrum secundum were studied in a semi-deciduous forest at Correspondence the Serra do Japi (SJ), and in the Atlantic rain forest of Picinguaba, both E. R. Pansarin, Departamento de Biologia natural reserves in the State of Sa˜o Paulo, southeastern Brazil. E. secundum Aplicada, Universidade Estadual Paulista, flowers all year round, with a flowering peak between September and FCAV, 14884-900, Jaboticabal, SP, Brazil. January. This species is either a lithophytic or terrestrial herb in the SJ, E-mail: [email protected] whereas, in Picinguaba, it grows mainly in disturbed areas along roadsides. E. secundum is pollinated by several species of diurnal Lepidoptera at both Editor study sites. In Picinguaba, where E. secundum is sympatric with E. fulgens M. Ayasse and both share the same pollinators, pollen transference between these two species was recorded. E. secundum is self-compatible but pollinator-depen- Received: 25 March 2007; Accepted: 22 May dent. It is inter-compatible with E. fulgens, producing fertile seeds. In con- 2007 trast to the population of the SJ, in the Picinguaba region, floral morphology is quite variable among plants and some individuals present doi:10.1111/j.1438-8677.2007.00025.x flowers with characteristics in-between both sympatric species, suggesting that natural hybridization occasionally occurs. -
00004-2005 ( .Pdf )
ESTE NÚMERO DE LANKESTERIANA FUE PUBLICADO GRACIAS AL GENTIL PATROCINIO DE LA FUNDACIÓN CHARLES H. LANKESTER ISSN 1409-3871 VOL. 5, No. 1 ABRIL 2005 Memoria de uno de los grandes de la orquideología mundial: Karlheinz Senghas (1928-2004) CARLOS O. MORALES 1 Análisis de la literatura sobre plantas medicinales en Costa Rica (1930-2001) MILDRED GARCÍA-GONZÁLEZ & CARLOS O. MORALES 3 El complejo de Campyloneurum angustifolium (Sw.) Fée (Polypodiaceae) en Costa Rica ALEXÁNDER FCO. ROJAS ALVARADO 41 Una nueva especie de Blechnum L. (Blechnaceae) en el neotrópico ALEXÁNDER FCO. ROJAS ALVARADO 49 Bryophytes of the Santa Elena Peninsula and Islas Murciélago, Guanacaste, Costa Rica, with special attention to neotropical dry forest habitats GREGORIO DAUPHIN L. & MICHAEL H. GRAYUM 53 Epicladium Small or Guarianthe Dressler & W.E. Higgins (Orchidaceae)? JOHN BECKNER 63 Chemical composition of the leaf oil of Peperomia hernandiifolia (Piperaceae) from Costa Rica JOSÉ F. CICCIÓ 69 Transfers to Epidendrum L. from Oerstedella Rchb. f. ERIC HÁGSATER & MIGUEL A. SOTO ARENAS 73 Continúa LA REVISTA CIENTÍFICA DEL JARDÍN BOTÁNICO LANKESTER UNIVERSIDAD DE COSTA RICA LANKESTERIANA Another new species of Stanhopea (Orchidaceae) from Peru RUDOLF JENNY 77 Una nueva especie y un nuevo registro de Drymonia (Gesneriaceae) en Costa Rica RICARDO KRIEBEL 81 Sobre un opus magnum de la orquideología neotropical: HÁGSATER, E. & SÁNCHEZ SALDAÑA, L. (EDS.). 2004. ICONES ORCHIDACEARUM, FASC. 7, THE GENUS EPIDENDRUM, PART 4, “A FOURTH CENTURY OF NEW SPECIES IN EPIDENDRUM” CARLOS O. MORALES 85 LANKESTERIANA LA REVISTA CIENTÍFICA DEL JARDÍN BOTÁNICO LANKESTER UNIVERSIDAD DE COSTA RICA Copyright © 2005 Jardín Botánico Lankester, Universidad de Costa Rica Fecha efectiva de publicación / Effective publication date: 30 de abril del 2005 Diagramación: Jardín Botánico Lankester Imprenta: Litografía Ediciones Sanabria S.A. -

Encyclia Fimbriata (Orchidaceae: Laeliinae), a New Large-Flowered Species from Bahia, Brazil
Phytotaxa 40: 26–40 (2012) ISSN 1179-3155 (print edition) www.mapress.com/phytotaxa/ Article PHYTOTAXA Copyright © 2012 Magnolia Press ISSN 1179-3163 (online edition) Encyclia fimbriata (Orchidaceae: Laeliinae), a new large-flowered species from Bahia, Brazil CLÁUDIA A. BASTOS1, CÁSSIO VAN DEN BERG1 & THIAGO E.C. MENEGUZZO2 1Universidade Estadual de Feira de Santana, Programa de Pós-graduação em Botânica. Av. Transnordestina, s.n., 44036-900, Feira de Santana, Bahia, Brazil; email: [email protected] 2 Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Rua Pacheco Leão, 915, Jardim Botânico, 22460-030, Rio de Janeiro, Rio de Janeiro, Brazil Abstract A new orchid species from Brazil, Encyclia fimbriata, is described and illustrated. It is known only from montane rain forest in southern Bahia. Flower morphology, especially the fringed midlobe of the labellum, crested callus and teeth of the clinandrium, distinguish it from any other species of the genus. Key words: Epiphytes, flora of Bahia, Neotropics Resumo Uma nova espécie de orquídea do Brasil, Encyclia fimbriata, é descrita e ilustrada. Esta é somente conhecida de floresta montana do sul da Bahia. A morfologia da flor, especialmente o lobo medial do labelo fimbriado, o calo sinuoso e os dentes do clinândrio, a diferenciam de todas as demais espécies do gênero. Palavras-chave: Epífitas, flora da Bahia, Neotrópico Introduction Encyclia is the second largest genus in the Neotropical subtribe Laeliinae, being surpassed only by Epidendrum (Dressler 1993, Pridgeon et al. 2003, Chase et al. 2004). Many Encyclia species have ornamental value due to their large and showy flowers. There are about 120 species occuring from Florida, Mexico and West Indies to Brazil and northern Argentina (Withner 1998, 2000, van den Berg & Carnevali F.-C. -

Encyclia Xcordistes Sauleda) from Colombia Is Described
ISSN 2325-4785 New World Orchidaceae – Nomenclatural Notes Nomenclatural Note – Issue No. 60 November 24, 2019 A Second New Natural Hybrid of Encyclia Hook. (Encyclia xcordistes Sauleda) From Colombia is Described. Ruben P. Sauleda 6442 S. W. 107 Ct., Miami, Fl. 33173 ABSTRACT A second natural hybrid of Encyclia Hook. has been discovered in Colombia and is here described. The genus Encyclia Hook. is represented in Mexico by approximately 46 species, in Brazil by approximately 44 species (WCSP, 2018) and in Cuba at present 41 species are known, all with a high number of endemics. In Colombia only 14 species of Encyclia have been recognized and one cleistogamous variety (Encyclia chloroleuca variety yotocoensis Sauleda and J. C. Uribe). Natural hybrids are common in the Orchidaceae especially in the genus Encyclia (Sauleda and Adams, 1984; 1990; Sauleda, 2016). However, in Colombia hybrids are rare, only one natural hybrid, Encyclia xortizii Sauleda and a highly polymorphic species, Encyclia replicata (Lindl. & Paxt.) Schltr. (Sauleda, 2016a) which appears to be a result of introgression with Encyclia ceratistes (Lindl.) Schltr. (Sauleda, 2016a) have been reported. Considering the high biodiversity of Colombia this low number of Encyclia species is unusual. In Colombia other genera like Epidendrum L. (503 species), Stelis Sw. (406 species), Pleurothallis R. Br. (205 species), Maxillaria Ruiz & Pav. (268 species) have species numbers in the hundreds. One would wonder why Colombia has only 14 species of Encyclia. A second natural hybrid of Encyclia with E. ceratistes has been discovered and is here described. Encyclia xcordistes Sauleda, hyb. nov. Encyclia cordigera (Kunth) Dressler X Encyclia ceratistes (Lindl.) Schltr. -
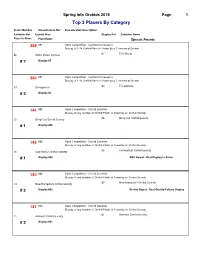
COS 2019 Show Ribbon Awards by Class
Spring Into Orchids 2019 Page 1 Top 3 Placers By Category Ticket Number Classification Ref Classification Description Exhibitor Ref Exhibit Area Display Ref Exhibitor Name Place In Class Plant Name Special Awards 359 001 Open Competition - Commercial Growers Display of 1-24 Orchid Plants in flower by a Commercial Grower 07 Eric Wang 06 White Plains Orchids # 2 Display #7 360 001 Open Competition - Commercial Growers Display of 1-24 Orchid Plants in flower by a Commercial Grower 03 Ecuagenera 37 Ecuagenera # 3 Display #3 144 003 Open Competition - Orchid Societies Display of any number of Orchid Plants in flower by an Orchid Society 06 Deep Cut Orchid Society 15 Deep Cut Orchid Society # 1 Display #06 145 003 Open Competition - Orchid Societies Display of any number of Orchid Plants in flower by an Orchid Society 08 Connecticut Orchid Society 16 Connecticut Orchid Society # 1 Display #08 AOS Award - Best Display in Show 140 003 Open Competition - Orchid Societies Display of any number of Orchid Plants in flower by an Orchid Society 05 New Hampshire Orchid Society 14 New Hampshire Orchid Society # 2 Display #05 Orchid Digest - Best Quality/Culture Display 141 003 Open Competition - Orchid Societies Display of any number of Orchid Plants in flower by an Orchid Society 01 Amherst Orchid Society 11 Amherst Orchid Society # 2 Display #01 Spring Into Orchids 2019 Page 2 Top 3 Placers By Category Ticket Number Classification Ref Classification Description Exhibitor Ref Exhibit Area Display Ref Exhibitor Name Place In Class Plant Name Special Awards 143 003 Open Competition - Orchid Societies Display of any number of Orchid Plants in flower by an Orchid Society 04 Cape & Islands Orchid Society 13 Cape & Islands Orchid Society # 2 Display #04 100 011 Cattleya Alliance(Laeliinae) Encyclia species 05 Chuck & Sue Andersen 10 New Hampshire Orchid Society # 1 Encyclia vitellina 67 011 Cattleya Alliance(Laeliinae) Encyclia species 07 Eric Wang 06 White Plains Orchids # 2 Enc. -

Epilist 1.0: a Global Checklist of Vascular Epiphytes
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2021 EpiList 1.0: a global checklist of vascular epiphytes Zotz, Gerhard ; Weigelt, Patrick ; Kessler, Michael ; Kreft, Holger ; Taylor, Amanda Abstract: Epiphytes make up roughly 10% of all vascular plant species globally and play important functional roles, especially in tropical forests. However, to date, there is no comprehensive list of vas- cular epiphyte species. Here, we present EpiList 1.0, the first global list of vascular epiphytes based on standardized definitions and taxonomy. We include obligate epiphytes, facultative epiphytes, and hemiepiphytes, as the latter share the vulnerable epiphytic stage as juveniles. Based on 978 references, the checklist includes >31,000 species of 79 plant families. Species names were standardized against World Flora Online for seed plants and against the World Ferns database for lycophytes and ferns. In cases of species missing from these databases, we used other databases (mostly World Checklist of Selected Plant Families). For all species, author names and IDs for World Flora Online entries are provided to facilitate the alignment with other plant databases, and to avoid ambiguities. EpiList 1.0 will be a rich source for synthetic studies in ecology, biogeography, and evolutionary biology as it offers, for the first time, a species‐level overview over all currently known vascular epiphytes. At the same time, the list represents work in progress: species descriptions of epiphytic taxa are ongoing and published life form information in floristic inventories and trait and distribution databases is often incomplete and sometimes evenwrong. -

Sistemática Y Evolución De Encyclia Hook
·>- POSGRADO EN CIENCIAS ~ BIOLÓGICAS CICY ) Centro de Investigación Científica de Yucatán, A.C. Posgrado en Ciencias Biológicas SISTEMÁTICA Y EVOLUCIÓN DE ENCYCLIA HOOK. (ORCHIDACEAE: LAELIINAE), CON ÉNFASIS EN MEGAMÉXICO 111 Tesis que presenta CARLOS LUIS LEOPARDI VERDE En opción al título de DOCTOR EN CIENCIAS (Ciencias Biológicas: Opción Recursos Naturales) Mérida, Yucatán, México Abril 2014 ( 1 CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C. POSGRADO EN CIENCIAS BIOLÓGICAS OSCJRA )0 f CENCIAS RECONOCIMIENTO S( JIOI ÚGIC A'- CICY Por medio de la presente, hago constar que el trabajo de tesis titulado "Sistemática y evo lución de Encyclia Hook. (Orchidaceae, Laeliinae), con énfasis en Megaméxico 111" fue realizado en los laboratorios de la Unidad de Recursos Naturales del Centro de Investiga ción Científica de Yucatán , A.C. bajo la dirección de los Drs. Germán Carnevali y Gustavo A. Romero, dentro de la opción Recursos Naturales, perteneciente al Programa de Pos grado en Ciencias Biológicas de este Centro. Atentamente, Coordinador de Docencia Centro de Investigación Científica de Yucatán, A.C. Mérida, Yucatán, México; a 26 de marzo de 2014 DECLARACIÓN DE PROPIEDAD Declaro que la información contenida en la sección de Materiales y Métodos Experimentales, los Resultados y Discusión de este documento, proviene de las actividades de experimen tación realizadas durante el período que se me asignó para desarrollar mi trabajo de tesis, en las Unidades y Laboratorios del Centro de Investigación Científica de Yucatán, A.C., y que a razón de lo anterior y en contraprestación de los servicios educativos o de apoyo que me fueron brindados, dicha información, en términos de la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, le pertenece patrimonialmente a dicho Centro de Investigación. -

Epidendrum L. (Orchidaceae, Epidendroideae) No Parque Nacional Da Chapada Dos Veadeiros, Estado De Goiás, Brasil
Artigo Hoehnea 47: e202020, 9 fig., 2020 http://dx.doi.org/10.1590/2236-8906-20/2020 Epidendrum L. (Orchidaceae, Epidendroideae) no Parque Nacional da Chapada dos Veadeiros, Estado de Goiás, Brasil Igor Soares dos Santos1,2 & Marcos José da Silva1 Recebido: 20.03.2020; aceito: 05.08.2020 Como citar: Santos, I.S. & Silva, M.J. 2020. Epidendrum L. (Orchidaceae, Epidendroideae) no Parque Nacional da Chapada dos Veadeiros, Estado de Goiás, Brasil. Hoehnea 47: e202020. http://dx.doi.org/10.1590/2236-8906-20/2020. RESUMO – (Epidendrum L. (Orchidaceae, Epidendroideae) no Parque Nacional da Chapada dos Veadeiros, Estado de Goiás, Brasil). Epidendrum L. é um dos maiores gêneros de Orchidaceae Juss. com 2.400 espécies neotropicais, 121 das quais presentes no Brasil, sendo 69 delas endêmicas. A taxonomia das espécies de Epidendrum na região Centro-Oeste é escassamente conhecida e vinculada a estudos florísticos sobre Orchidaceae. É apresentado o tratamento taxonômico às espécies de Epidendrum ocorrentes no Parque Nacional da Chapada dos Veadeiros, uma das Áreas de Preservação Permanentes mais importantes do Brasil. Foram encontradas seis espécies: E. avicula Lindl., E. campacci Hágsater & L. Sánchez, E. dendrobioides Thunb., E. nocturnum Jacq., E. rothii A.D. Hawkes e E. secundum Jacq., crescendo como epífitas, terrícolas e rupícolas em distintas fitofisionomias. As espécies seguem descritas e ilustradas, comentadas quanto a distribuição geográfica, relações morfológicas, fenologia, bem como contrastadas por meio de uma chave dicotômica e alocadas em grupos informais reconhecidos para o gênero. Palavras-chave: Cerrado, diversidade, flora, orquídeas, Taxonomia ABSTRACT – (Epidendrum L. (Orchidaceae, Epidendroideae) in the Parque Nacional da Chapada dos Veadeiros, Goiás State, Brazil).