JNS-3AP-Ibo-Pl1fin.Qrk
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Floor Plate and Netrin-1 Are Involved in the Migration and Survival of Inferior Olivary Neurons
The Journal of Neuroscience, June 1, 1999, 19(11):4407–4420 Floor Plate and Netrin-1 Are Involved in the Migration and Survival of Inferior Olivary Neurons Evelyne Bloch-Gallego,1 Fre´de´ ric Ezan,1 Marc Tessier-Lavigne,2 and Constantino Sotelo1 1Institut National de la Sante´ et de la Recherche Me´ dicale U106, Hoˆ pital de la Salpeˆ trie` re, 75013 Paris, France, and 2Howard Hughes Medical Institute, Department of Anatomy, University of California at San Francisco, San Francisco, California 94143 During their circumferential migration, the nuclei of inferior oli- However, axons of the remaining olivary cell bodies located in vary neurons translocate within their axons until they reach the the vicinity of the floor plate still succeed in entering their target, floor plate where they stop, although their axons have already the cerebellum, but they establish an ipsilateral projection in- crossed the midline to project to the contralateral cerebellum. stead of the normal contralateral projection. In vitro experi- Signals released by the floor plate, including netrin-1, have ments involving ablations of the midline show a fusion of the been implicated in promoting axonal growth and chemoattrac- two olivary masses normally located on either side of the tion during axonal pathfinding in different midline crossing sys- ventral midline, suggesting that the floor plate may function as tems. In the present study, we report experiments that strongly a specific stop signal for inferior olivary neurons. These results suggest that the floor plate could also be involved in the establish a requirement for netrin-1 in the migration of inferior migration of inferior olivary neurons. -

Basal Ganglia & Cerebellum
1/2/2019 This power point is made available as an educational resource or study aid for your use only. This presentation may not be duplicated for others and should not be redistributed or posted anywhere on the internet or on any personal websites. Your use of this resource is with the acknowledgment and acceptance of those restrictions. Basal Ganglia & Cerebellum – a quick overview MHD-Neuroanatomy – Neuroscience Block Gregory Gruener, MD, MBA, MHPE Vice Dean for Education, SSOM Professor, Department of Neurology LUHS a member of Trinity Health Outcomes you want to accomplish Basal ganglia review Define and identify the major divisions of the basal ganglia List the major basal ganglia functional loops and roles List the components of the basal ganglia functional “circuitry” and associated neurotransmitters Describe the direct and indirect motor pathways and relevance/role of the substantia nigra compacta 1 1/2/2019 Basal Ganglia Terminology Striatum Caudate nucleus Nucleus accumbens Putamen Globus pallidus (pallidum) internal segment (GPi) external segment (GPe) Subthalamic nucleus Substantia nigra compact part (SNc) reticular part (SNr) Basal ganglia “circuitry” • BG have no major outputs to LMNs – Influence LMNs via the cerebral cortex • Input to striatum from cortex is excitatory – Glutamate is the neurotransmitter • Principal output from BG is via GPi + SNr – Output to thalamus, GABA is the neurotransmitter • Thalamocortical projections are excitatory – Concerned with motor “intention” • Balance of excitatory & inhibitory inputs to striatum, determine whether thalamus is suppressed BG circuits are parallel loops • Motor loop – Concerned with learned movements • Cognitive loop – Concerned with motor “intention” • Limbic loop – Emotional aspects of movements • Oculomotor loop – Concerned with voluntary saccades (fast eye-movements) 2 1/2/2019 Basal ganglia “circuitry” Cortex Striatum Thalamus GPi + SNr Nolte. -

Molecular Plasticity of Adult Bergmann Fibers Is Associated with Radial Migration of Grafted Purkinje Cells
The Journal of Neuroscience, January 1994, 14(i): 124-133 Molecular Plasticity of Adult Bergmann Fibers Is Associated with Radial Migration of Grafted Purkinje Cells Constantino Sotelo,’ Rosa-Magda Alvarado-Mallart,i Monique Frain,2 and Muriel Verner ‘Neuromorphologie: Dkveloppement, Evolution, INSERM U. 106, HBpital de la Salpktrikre, 75651 Paris Cedex 13, France, *Biologic Molkulaire du Dkveloppement, INSERM U. 368 Ecole Normale Supkrieure, 75230 Paris Cedex, France, and 31nstitut Cochin de Gbnbtique Molkulaire, CJF 9003 INSERM, 75014 Paris, France Embryonic Purkinje cells (PCs) from cerebellar primordia adult cerebellum and become functionally integrated into the grafted in adult pcd mutant cerebellum replace missing PCs synaptic circuitry of the cerebellarcortex of the host (Sotelo and of the host, and become synaptically integrated into the de- Alvarado-Mallart, 1991). The cerebellar mutant mouse strains fective cerebellar circuit. This process of neuronal replace- .LJC~,nr, and Lc, in which Purkinje cells (PCs) die (Sidman and ment starts with the invasion of grafted PCs into the host Green, 1970; Mullen et al., 1976; Caddy and Biscoe, 1979; cerebellum, and their radial migration through its molecular Wassef et al., 1987; Dumesnil-Bousez and Sotelo, 1992), have layer. The present study is aimed at determining whether provided models for studying graft integration (Sotelo and Al- the glial axes for this migration are embryonic radial glial varado-Mallart, 1986, 1987a, 1991, 1992; Dumesnil-Bousezand cells that comigrate with the grafted PCs, or adult Bergmann Sotelo, 1993). Penetration of grafted PCs into the cerebellum fibers of the host, transiently reexpressing the molecular of mutant hosts begins 3-4 d after grafting (DAG 3-4) by tan- cues needed for their guidance of the migration. -
L4-Physiology of Motor Tracts.Pdf
: chapter 55 page 667 Objectives (1) Describe the upper and lower motor neurons. (2) Understand the pathway of Pyramidal tracts (Corticospinal & corticobulbar tracts). (3) Understand the lateral and ventral corticospinal tracts. (4) Explain functional role of corticospinal & corticobulbar tracts. (5) Describe the Extrapyramidal tracts as Rubrospinal, Vestibulospinal, Reticulospinal and Tectspinal Tracts. The name of the tract indicate its pathway, for example Corticobulbar : Terms: - cortico: cerebral cortex. Decustation: crossing. - Bulbar: brainstem. Ipsilateral : same side. *So it starts at cerebral cortex and Contralateral: opposite side. terminate at the brainstem. CNS influence the activity of skeletal muscle through two set of neurons : 1- Upper motor neurons (UMN) 2- lower motor neuron (LMN) They are neurons of motor cortex & their axons that pass to brain stem and They are Spinal motor neurons in the spinal spinal cord to activate: cord & cranial motor neurons in the brain • cranial motor neurons (in brainstem) stem which innervate muscles directly. • spinal motor neurons (in spinal cord) - These are the only neurons that innervate - Upper motor neurons (UMN) are the skeletal muscle fibers, they function as responsible for conveying impulses for the final common pathway, the final link voluntary motor activity through between the CNS and skeletal muscles. descending motor pathways that make up by the upper motor neurons. Lower motor neurons are classified based on the type of muscle fiber the innervate: There are two UMN Systems through which 1- alpha motor neurons (UMN) control (LMN): 2- gamma motor neurons 1- Pyramidal system (corticospinal tracts ). 2- Extrapyramidal system The activity of the lower motor neuron (LMN, spinal or cranial) is influenced by: 1. -

Neurochemical and Structural Organization of the Principal Nucleus of the Inferior Olive in the Human
THE ANATOMICAL RECORD 294:1198–1216 (2011) Neurochemical and Structural Organization of the Principal Nucleus of the Inferior Olive in the Human JOAN S. BAIZER,1* CHET C. SHERWOOD,2 PATRICK R. HOF,3 4 5 SANDRA F. WITELSON, AND FAHAD SULTAN 1Department of Physiology and Biophysics, University at Buffalo, Buffalo, New York 2Department of Anthropology, The George Washington University, Washington, District of Columbia 3Department of Neuroscience and Friedman Brain Institute, Mount Sinai School of Medicine, New York, New York 4Department of Psychiatry & Behavioural Neurosciences, Michael G. DeGroote School of Medicine, McMaster University, Hamilton, Ontario, Canada L8N 3Z5 5Department of Cognitive Neurology, University of Tu¨ bingen, Tu¨ bingen, Germany ABSTRACT The inferior olive (IO) is the sole source of the climbing fibers that innervate the Purkinje cells of the cerebellar cortex. The IO comprises several subdivisions, the dorsal accessory olive (DAO), medial accessory olive (MAO), and principal nuclei of the IO (IOpr); the relative sizes of these subnuclei vary among species. In human, there is an expansion of the cerebellar hemispheres and a corresponding expansion of the IOpr. We have examined the structural and neurochemical organization of the human IOpr, using sections stained with cresyl violet (CV) or immuno- stained for the calcium-binding proteins calbindin (CB), calretinin (CR), and parvalbumin (PV), the synthetic enzyme for nitric oxide (nNOS), and nonphosphorylated neurofilament protein (NPNFP). We found qualitative differences in the folding patterns of the IOpr among individuals and between the two sides of the brainstem. Quantification of IOpr volumes and indices of folding complexity, however, did not reveal consistent left–right differences in either parameter. -

Cerebellum and Inferior Olive
Cerebellum and Inferior Olivary Nucleus Spinocerebellum • Somatotopically organised (vermis controls axial musculature; intermediate hemisphere controls limb musculature) • Control of body musculature • Inputs… Vermis receives somatosensory information (mainly from the trunk) via the spinocerebellar tracts and from the spinal nucleus of V. It receives a direct projection from the primary sensory neurons of the vestibular labyrinth, and also visual and auditory input from brain stem nuclei. • Intermediate hemisphere receives somatosensory information (mainly from the limbs) via the spinocerebellar tracts (the dorsal spinocerebellar tract, from Clarke’s nucleus of the lower limb, and the cuneocerebellar tract, from the accessory cu- neate nucleus of the upper limb, carry information from muscle spindle afferents; both enter via the ipsilateral inferior cerebellar peduncle). • An internal feedback signal arrives via the ventral spinocerebellar tract (lower limb) and rostral spinocerebellar tract (upper limb). (Ventral s.t. decussates in the spinal cord and enters via the superior cerebellar peduncle, but some fibres re-cross in the cerebellum; rostral s.t. is an ipsilateral pathway and enters via sup. & inf. cerebellar peduncles.) • Outputs to fastigial nucleus, which projects to the medial descending systems: (1) reticulospinal tract [? n. reticularis teg- menti pontis and prepositus hypoglossi?]; (2) vestibulospinal tract [lateral and descending vestibular nn.]; and (3) an as- cending projection to VL thalamus [Å cells of origin of the ventral corticospinal tract]; (4) reticular grey of the midbrain [=periaqueductal?]; (5) inferior olive [medial accessory, MAO]. • … and interposed nuclei, which project to the lateral descending systems: (1) magnocellular portion of red nucleus [Å ru- brospinal tract]; (2) VL thalamus [Å motor cx which gives rise to lateral corticospinal tract]; (3) reticular nucleus of the pontine tegmentum; (4) inferior olive [dorsal accessory, DAO]; (5) spinal cord intermediate grey. -

Cerebellar Granule Cells in Culture
Proc. Nati. Acad. Sci. USA Vol. 83, pp. 4957-4961, July 1986 Neurobiology Cerebellar granule cells in culture: Monosynaptic connections with Purkinje cells and ionic currents (excitatory postsynaptic potential/patch-clamp) ToMoo HIRANO, YOSHIHIRo KUBO, AND MICHAEL M. WU Department of Neurobiology, Institute of Brain Research, School of Medicine, University of Tokyo, Tokyo, Japan Communicated by S. Hagiwara, March 6, 1986 ABSTRACT Electrophysiological properties of cerebellar tissue was dissociated by triturating with a fire-polished granule cells and synapses between granule and Purkinje cells Pasteur pipette in Ca-free Hanks' balanced salt solution were studied in dissociated cultures. Electrophysiological prop- containing 0.05% DNase and 12 mM MgSO4. The cells were erties of neurons and synapses in the mammalian central centrifuged at 150 x g at 40C and the pelleted cells were nervous system are best studied in dissociated cell cultures resuspended at a concentration of about 106 cells per ml in a because of good target cell visibility, control over the contents defined medium (9). One milliliter ofthe cell suspension from of the extracellular solution, and the feasibility of whole-cell newborn rats was plated first in a Petri dish (3.5 cm in patch electrode recording, which has been a powerful tech- diameter) containing several pieces ofheat-sterilized, poly(L- nique in analyzing biophysical properties of ionic channels in lysine)-coated coverslips, and then 1 ml of fetal cell suspen- small cells. We have applied this whole-cell recording technique sion was added. One-half of the culture medium was ex- to cultured cerebellar granule cells whose electrophysiological changed with fresh medium once a week. -
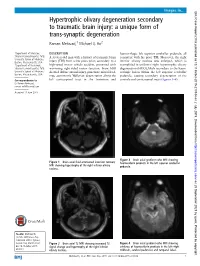
Hypertrophic Olivary Degeneration Secondary to Traumatic Brain Injury: a Unique Form of Trans-Synaptic Degeneration Raman Mehrzad,1 Michael G Ho2
… Images in BMJ Case Reports: first published as 10.1136/bcr-2015-210334 on 2 July 2015. Downloaded from Hypertrophic olivary degeneration secondary to traumatic brain injury: a unique form of trans-synaptic degeneration Raman Mehrzad,1 Michael G Ho2 1Department of Medicine, DESCRIPTION haemorrhagic left superior cerebellar peduncle, all Steward Carney Hospital, Tufts A 33-year-old man with a history of traumatic brain consistent with his prior TBI. Moreover, the right University School of Medicine, Boston, Massachusetts, USA injury (TBI) from a few years prior, secondary to a inferior olivary nucleus was enlarged, which is 2Department of Neurology, high-speed motor vehicle accident, presented with exemplified in unilateral right hypertrophic olivary Steward Carney Hospital, Tufts worsening right-sided motor function. Brain MRI degeneration (HOD), likely secondary to the haem- University School of Medicine, showed diffuse axonal injury, punctuate microbleed- orrhagic lesion within the left superior cerebellar Boston, Massachusetts, USA ings, asymmetric Wallerian degeneration along the peduncle, causing secondary degeneration of the fi – Correspondence to left corticospinal tract in the brainstem and contralateral corticospinal tracts ( gures 1 6). Dr Raman Mehrzad, [email protected] Accepted 11 June 2015 http://casereports.bmj.com/ fl Figure 3 Brain axial gradient echo MRI showing Figure 1 Brain axial uid-attenuated inversion recovery haemosiderin products in the left superior cerebellar MRI showing hypertrophy of the right inferior olivary peduncle. nucleus. on 25 September 2021 by guest. Protected copyright. To cite: Mehrzad R, Ho MG. BMJ Case Rep Published online: [please include Day Month Year] Figure 2 Brain axial T2 MRI showing increased T2 Figure 4 Brain axial gradient echo MRI showing doi:10.1136/bcr-2015- signal change and hypertrophy of the right inferior evidence of haemosiderin products in the left>right 210334 olivary nucleus. -

FIRST PROOF Cerebellum
Article Number : EONS : 0736 GROSS ANATOMY Cerebellum Cortex The cerebellar cortex is an extensive three-layered sheet with a surface approximately 15 cm laterally THE HUMAN CEREBELLUM (‘‘little brain’’) is a and 180 cm rostrocaudally but densely folded around significant part of the central nervous system both three pairs of nuclei. The cortex is divided into three in size and in neural structure. It occupies approxi- transverse lobes: Anterior and posterior lobes are mately one-tenth of the cranial cavity, sitting astride separated by the primary fissure, and the smaller the brainstem, beneath the occipital cortex, and flocculonodular lobe is separated by the poster- contains more neurons than the whole of the cerebral olateral fissure (Fig. 1). The anterior and posterior cortex. It consists of an extensive cortical sheet, lobes are folded into a number of lobules and further densely folded around three pairs of nuclei. The folded into a series of folia. This transverse organiza- cortex contains only five main neural cell types and is tion is then divided at right angles into broad one of the most regular and uniform structures in the longitudinal regions. The central vermis, named for central nervous system (CNS), with an orthogonal its worm-like appearance, is most obvious in the ‘‘crystalline’’ organization. Major connections are posterior lobe. On either side is the paravermal or made to and from the spinal cord, brainstem, and intermediate cortex, which merges into the lateral sensorimotor areas of the cerebral cortex. hemispheres. The most common causes of damage to the cerebellum are stroke, tumors, or multiple sclerosis. -

Three-Dimensional Comparison of Ultrastructural Characteristics at Depressing and Facilitating Synapses Onto Cerebellar Purkinje Cells
The Journal of Neuroscience, September 1, 2001, 21(17):6666–6672 Three-Dimensional Comparison of Ultrastructural Characteristics at Depressing and Facilitating Synapses onto Cerebellar Purkinje Cells Matthew A. Xu-Friedman,1 Kristen M. Harris,2 and Wade G. Regehr1 1Department of Neurobiology, Harvard Medical School, Boston, Massachusetts 02115, and 2Department of Biology, Boston University, Boston, Massachusetts 02215 Cerebellar Purkinje cells receive two distinctive types of exci- vesicles per release site to test whether this number determines tatory inputs. Climbing fiber (CF) synapses have a high proba- the probability of release and synaptic plasticity. PF and CF bility of release and show paired-pulse depression (PPD), synapses had the same number of anatomically docked vesi- whereas parallel fiber (PF) synapses facilitate and have a low cles (7–8). The number of docked vesicles at the CF does not probability of release. We examined both types of synapses support a simple model of PPD in which release of a single using serial electron microscopic reconstructions in 15-d-old vesicle during the first pulse depletes the anatomically docked rats to look for anatomical correlates of these differences. PF vesicle pool at a synapse. Alternatively, only a fraction of ana- and CF synapses were distinguishable by their overall ultra- tomically docked vesicles may be release ready, or PPD could structural organization. There were differences between PF and result from multivesicular release at each site. Similarities in the CF synapses in how many release sites were within 1 mofa number of docked vesicles for PF and CF synapses indicate mitochondrion (67 vs 84%) and in the degree of astrocytic that differences in probability of release are unrelated to the ensheathment (67 vs 94%). -

Profound Functional and Molecular Diversity of Mitochondria Revealed by Cell Type-Specific Profiling in Vivo
bioRxiv preprint doi: https://doi.org/10.1101/403774; this version posted August 29, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Profound functional and molecular diversity of mitochondria revealed by cell type-specific profiling in vivo Authors: Caroline Fecher1,2,3,#, Laura Trovò1,2,#, Stephan A. Müller2,4, Nicolas Snaidero1,2, Jennifer Wettmarshausen5,6, Sylvia Heink7, Oskar Ortiz8,$, Ingrid Wagner9, Ralf Kühn8,$$, Jana Hartmann10, Rosa Maria Karl10, Arthur Konnerth10,11,12, Thomas Korn7,11, Wolfgang Wurst2,8,11,13, Doron Merkler9, Stefan F. Lichtenthaler2,4,11, Fabiana Perocchi5,6,11 and Thomas Misgeld1,2,11,12,* Affiliations: 1 Institute of Neuronal Cell Biology, Technical University of Munich, Germany 2 German Center for Neurodegenerative Diseases, Munich, Germany 3 Graduate School of Systemic Neuroscience, Ludwig-Maximilians University, Munich, Germany 4 Neuroproteomics, School of Medicine, Klinikum rechts der Isar, and Institute for Advanced Study, Technical University of Munich, Germany 5 Gene Center Munich, Ludwig-Maximilians University, Munich, Germany 6 Institute for Diabetes and Obesity, Helmholtz Zentrum München, Munich, Germany 7 Experimental Neuroimmunology, Klinikum rechts der Isar, Technical University of Munich, Germany 8 Institute of Developmental Genetics, Helmholtz Zentrum München, Munich, Germany 9 Department of -

Holmes Tremor in Association with Bilateral Hypertrophic Olivary Degeneration and Palatal Tremor
Arq Neuropsiquiatr 2003;61(2-B):473-477 HOLMES TREMOR IN ASSOCIATION WITH BILATERAL HYPERTROPHIC OLIVARY DEGENERATION AND PALATAL TREMOR CHRONOLOGICAL CONSIDERATIONS Case report Carlos R.M. Rieder1, Ricardo Gurgel Rebouças2, Marcelo Paglioli Ferreira3 ABSTRACT - Hypertrophic olivary degeneration (HOD) is a rare type of neuronal degeneration involving the dento-rubro-olivary pathway and presents clinically as palatal tremor. We present a 48 year old male patient who developed Holmes’ tremor and bilateral HOD five months after brainstem hemorrhage. The severe rest tremor was refractory to pharmacotherapy and botulinum toxin injections, but was markedly reduced after thalamotomy. Magnetic resonance imaging permitted visualization of HOD, which appeared as a characteristic high signal intensity in the inferior olivary nuclei on T2- and proton-density-weighted images. Enlargement of the inferior olivary nuclei was also noted. Palatal tremor was absent in that moment and appears about two months later. The delayed-onset between insult and tremor following structural lesions of the brain suggest that compensatory or secondary changes in nervous system function must contribute to tremor genesis. The literature and imaging findings of this uncommon condition are reviewed. KEY WORDS: rubral tremor, midbrain tremor, Holmes’ tremor, myorhythmia, palatal myoclonus. Tremor de Holmes em associação com degeneração olivar hipertrófica bilateral e tremor palatal: considerações cronológicas. Relato de caso RESUMO - Degeneração olivar hipertrófica (DOH) é um tipo raro de degeneração neuronal envolvendo o trato dento-rubro-olivar e se apresenta clinicamente como tremor palatal. Relatamos o caso de um homem de 48 anos que desenvolveu tremor de Holmes e DOH bilateral cinco meses após hemorragia em tronco encefálico.