Original Article / Artigo Original
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Qt9z7703dj.Pdf
UC San Diego UC San Diego Previously Published Works Title Phylogeny and biogeography of a shallow water fish clade (Teleostei: Blenniiformes) Permalink https://escholarship.org/uc/item/9z7703dj Journal BMC Evolutionary Biology, 13(1) ISSN 1471-2148 Authors Lin, Hsiu-Chin Hastings, Philip A Publication Date 2013-09-25 DOI http://dx.doi.org/10.1186/1471-2148-13-210 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Lin and Hastings BMC Evolutionary Biology 2013, 13:210 http://www.biomedcentral.com/1471-2148/13/210 RESEARCH ARTICLE Open Access Phylogeny and biogeography of a shallow water fish clade (Teleostei: Blenniiformes) Hsiu-Chin Lin1,2* and Philip A Hastings1 Abstract Background: The Blenniiformes comprises six families, 151 genera and nearly 900 species of small teleost fishes closely associated with coastal benthic habitats. They provide an unparalleled opportunity for studying marine biogeography because they include the globally distributed families Tripterygiidae (triplefin blennies) and Blenniidae (combtooth blennies), the temperate Clinidae (kelp blennies), and three largely Neotropical families (Labrisomidae, Chaenopsidae, and Dactyloscopidae). However, interpretation of these distributional patterns has been hindered by largely unresolved inter-familial relationships and the lack of evidence of monophyly of the Labrisomidae. Results: We explored the phylogenetic relationships of the Blenniiformes based on one mitochondrial (COI) and four nuclear (TMO-4C4, RAG1, Rhodopsin, and Histone H3) loci for 150 blenniiform species, and representative outgroups (Gobiesocidae, Opistognathidae and Grammatidae). According to the consensus of Bayesian Inference, Maximum Likelihood, and Maximum Parsimony analyses, the monophyly of the Blenniiformes and the Tripterygiidae, Blenniidae, Clinidae, and Dactyloscopidae is supported. -

Coastal and Marine Ecological Classification Standard (2012)
FGDC-STD-018-2012 Coastal and Marine Ecological Classification Standard Marine and Coastal Spatial Data Subcommittee Federal Geographic Data Committee June, 2012 Federal Geographic Data Committee FGDC-STD-018-2012 Coastal and Marine Ecological Classification Standard, June 2012 ______________________________________________________________________________________ CONTENTS PAGE 1. Introduction ..................................................................................................................... 1 1.1 Objectives ................................................................................................................ 1 1.2 Need ......................................................................................................................... 2 1.3 Scope ........................................................................................................................ 2 1.4 Application ............................................................................................................... 3 1.5 Relationship to Previous FGDC Standards .............................................................. 4 1.6 Development Procedures ......................................................................................... 5 1.7 Guiding Principles ................................................................................................... 7 1.7.1 Build a Scientifically Sound Ecological Classification .................................... 7 1.7.2 Meet the Needs of a Wide Range of Users ...................................................... -

Community Structure of Reef Fishes on a Remote Oceanic Island
CSIRO PUBLISHING Marine and Freshwater Research http://dx.doi.org/10.1071/MF14150 Community structure of reef fishes on a remote oceanic island (St Peter and St Paul’s Archipelago, equatorial Atlantic): the relative influence of abiotic and biotic variables Osmar J. LuizA,G, Thiago C. MendesB, Diego R. BarnecheA, Carlos G. W. FerreiraC, Ramon NoguchiD, Roberto C. Villac¸aB, Carlos A. RangelE, Joa˜o L. GaspariniF and Carlos E. L. FerreiraB ADepartment of Biological Sciences, Macquarie University, Sydney, NSW 2109, Australia. BDepartamento de Biologia Marinha, Universidade Federal Fluminense, Nitero´i, RJ, 24001-970, Brazil. CDepartamento de Oceanografia, Instituto de Estudos do Mar Almirante Paulo Moreira, Arraial do Cabo, RJ, 28930-000, Brazil. DPrograma de Po´s Graduac¸a˜o em Ecologia, Universidade Federal de Rio de Janeiro, Rio de Janeiro, RJ, 68020, Brazil. EProjeto Ilhas do Rio, Instituto Mar Adentro, Rio de Janeiro, RJ, 22031-071, Brazil. FDepartamento de Oceanografia e Ecologia, Universidade Federal do Espı´rito Santo, Vito´ria, ES, Brazil. GCorresponding author. Email: [email protected] Abstract. This study investigates the reef fish community structure of the world’s smallest remote tropical island, the St Peter and St Paul’s Archipelago, in the equatorial Atlantic. The interplay between isolation, high endemism and low species richness makes the St Peter and St Paul’s Archipelago ecologically simpler than larger and highly connected shelf reef systems, making it an important natural laboratory for ecology and biogeography, particularly with respect to the effects of abiotic and biotic factors, and the functional organisation of such a depauperate community. Boosted regression trees were used to associate density, biomass and diversity of reef fishes with six abiotic and biotic variables, considering the community both as a whole and segregated into seven trophic groups. -

The Morphology and Evolution of Tooth Replacement in the Combtooth Blennies
The morphology and evolution of tooth replacement in the combtooth blennies (Ovalentaria: Blenniidae) A THESIS SUBMITTED TO THE FACULTY OF THE UNIVERSITY OF MINNESOTA BY Keiffer Logan Williams IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE Andrew M. Simons July 2020 ©Keiffer Logan Williams 2020 i ACKNOWLEDGEMENTS I thank my adviser, Andrew Simons, for mentoring me as a student in his lab. His mentorship, kindness, and thoughtful feedback/advice on my writing and research ideas have pushed me to become a more organized and disciplined thinker. I also like to thank my committee: Sharon Jansa, David Fox, and Kory Evans for feedback on my thesis and during committee meetings. An additional thank you to Kory, for taking me under his wing on the #backdattwrasseup project. Thanks to current and past members of the Simons lab/office space: Josh Egan, Sean Keogh, Tyler Imfeld, and Peter Hundt. I’ve enjoyed the thoughtful discussions, feedback on my writing, and happy hours over the past several years. Thanks also to the undergraduate workers in the Simons lab who assisted with various aspects of my work: Andrew Ching and Edward Hicks for helping with histology, and Alex Franzen and Claire Rude for making my terms as curatorial assistant all the easier. In addition, thank you to Kate Bemis and Karly Cohen for conducting a workshop on histology to collect data for this research, and for thoughtful conversations and ideas relating to this thesis. Thanks also to the University of Guam and Laurie Raymundo for hosting me as a student to conduct fieldwork for this research. -

Atividade De Limpeza E Clientes De Elacatinus Figaro (Pisces: Gobiidae) Nos Recifes De Coral Dos Parrachos De Muriú, Nordeste Do Brasil
Atividade de limpeza e clientes de Elacatinus figaro (Pisces: Gobiidae) nos recifes de coral dos Parrachos de Muriú, Nordeste do Brasil Campos, C.E.C. & Sá-Oliveira, J.C. Biota Neotrop. 2011, 11(1): 47-52. On line version of this paper is available from: http://www.biotaneotropica.org.br/v11n1/en/abstract?article+bn01011012011 A versão on-line completa deste artigo está disponível em: http://www.biotaneotropica.org.br/v11n1/pt/abstract?article+bn01011012011 Received/ Recebido em 30/06/2010 - Revised/ Versão reformulada recebida em 14/12/2010 - Accepted/ Publicado em 11/01/2011 ISSN 1676-0603 (on-line) Biota Neotropica is an electronic, peer-reviewed journal edited by the Program BIOTA/FAPESP: The Virtual Institute of Biodiversity. This journal’s aim is to disseminate the results of original research work, associated or not to the program, concerned with characterization, conservation and sustainable use of biodiversity within the Neotropical region. Biota Neotropica é uma revista do Programa BIOTA/FAPESP - O Instituto Virtual da Biodiversidade, que publica resultados de pesquisa original, vinculada ou não ao programa, que abordem a temática caracterização, conservação e uso sustentável da biodiversidade na região Neotropical. Biota Neotropica is an eletronic journal which is available free at the following site http://www.biotaneotropica.org.br A Biota Neotropica é uma revista eletrônica e está integral e gratuitamente disponível no endereço http://www.biotaneotropica.org.br Biota Neotrop., vol. 11, no. 1 Atividade de limpeza e clientes de Elacatinus figaro (Pisces: Gobiidae) nos recifes de coral dos Parrachos de Muriú, Nordeste do Brasil Carlos Eduardo Costa Campos1,2 & Júlio César Sá-Oliveira1 1 Laboratório de Zoologia, Departamento de Ciências Biológicas, Universidade Federal do Amapá – UNIFAP, Rod. -

Herbivory by the Dusky Damselfish Stegastes Fuscus (Cuvier, 1830)
Journal of Experimental Marine Biology and Ecology, L 229 (1998) 241±264 Herbivory by the Dusky Damsel®sh Stegastes fuscus (Cuvier, 1830) in a tropical rocky shore: effects on the benthic community Carlos Eduardo L. Ferreiraa,b,* , Jose Eduardo A. GoncËalves b , Ricardo Coutinhoba , Alberto C. Peret aDep. de Hidrobiologia, Universidade Federal de SaoÄÄ Carlos, Sao Carlos, S.P., Cep:13560 000, Brazil bDept. de Biologia, Instituto de Estudos do Mar Almirante Paulo Moreira, Arraial do Cabo, R.J., Cep: 28930 000, Brazil Received 12 September 1995; received in revised form 31 March 1997; accepted 13 March 1998 Abstract Experiments were carried out on rocky shores at Arraial do Cabo (Southeast Brazil) to evaluate how the dusky damsel®sh, Stegastes fuscus (Cuvier, 1830) affects the benthic community structure. Cage exclusion showed that S. fuscus strongly in¯uences the algal community in its territories, keeping it in an early succession stage and preventing dominance by Jania spp. Diversity and biomass of the epilithic algal community (EAC) were higher inside territories than | 2 outside. These dense mats hold a diverse and abundant cryptofauna (5 72 ind/100 cm ) that was signi®cantly higher inside territories. Algae comprise 70% of the S. fuscus diet, with the remaining 30% composed of animal material. The ®sh feeds selectively mainly on red ®lamentous algae, such as Polysiphonia spp., Ceramium spp. and Centroceras clavulatum, albeit it also ingests a great amount of calcareous algae (25% of total algae). Total assimilation and nitrogen assimilation were low in S. fuscus. Gut contents turnover varied from 3.7 in summer to 4.1 in winter. -

Taverampe2018.Pdf
Molecular Phylogenetics and Evolution 121 (2018) 212–223 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Multilocus phylogeny, divergence times, and a major role for the benthic-to- T pelagic axis in the diversification of grunts (Haemulidae) ⁎ Jose Taveraa,b, , Arturo Acero P.c, Peter C. Wainwrightb a Departamento de Biología, Universidad del Valle, Cali, Colombia b Department of Evolution and Ecology, University of California, Davis, CA 95616, United States c Instituto de Estudios en Ciencias del Mar, CECIMAR, Universidad Nacional de Colombia sede Caribe, El Rodadero, Santa Marta, Colombia ARTICLE INFO ABSTRACT Keywords: We present a phylogenetic analysis with divergence time estimates, and an ecomorphological assessment of the Percomorpharia role of the benthic-to-pelagic axis of diversification in the history of haemulid fishes. Phylogenetic analyses were Fish performed on 97 grunt species based on sequence data collected from seven loci. Divergence time estimation Functional traits indicates that Haemulidae originated during the mid Eocene (54.7–42.3 Ma) but that the major lineages were Morphospace formed during the mid-Oligocene 30–25 Ma. We propose a new classification that reflects the phylogenetic Macroevolution history of grunts. Overall the pattern of morphological and functional diversification in grunts appears to be Zooplanktivore strongly linked with feeding ecology. Feeding traits and the first principal component of body shape strongly separate species that feed in benthic and pelagic habitats. The benthic-to-pelagic axis has been the major axis of ecomorphological diversification in this important group of tropical shoreline fishes, with about 13 transitions between feeding habitats that have had major consequences for head and body morphology. -

Universidade Federal Do Estado Do Rio De Janeiro Instituto De Biociências
1 UNIVERSIDADE FEDERAL DO ESTADO DO RIO DE JANEIRO INSTITUTO DE BIOCIÊNCIAS COMPORTAMENTO TERRITORIALISTA DO Stegastes fuscus (Cuvier, 1830) E A ASSEMBLEIA DE PEIXES RECIFAIS EM COSTÃO ROCHOSO TROPICAL: ANÁLISE DE SENSIBILIDADE Victor Bastos Teixeira Lupinacci Rio de Janeiro 2019 2 Victor Bastos Teixeira Lupinacci COMPORTAMENTO TERRITORIALISTA DO Stegastes fuscus (Cuvier, 1830) E A ASSEMBLEIA DE PEIXES RECIFAIS EM COSTÃO ROCHOSO TROPICAL: ANÁLISE DE SENSIBILIDADE Monografia do Trabalho de Conclusão de Curso apresentada ao Instituto de Biociências da Universidade Federal do Estado do Rio de Janeiro, como parte dos requisitos à obtenção do título de Bacharel em Ciências Ambientais. Orientador: Prof. Dr. Rafael da Rocha Fortes Rio de Janeiro 2019 3 LUPINACCI, Victor COMPORTAMENTO TERRITORIALISTA DO Stegastes fuscus (Cuvier, 1830) E A ASSEMBLEIA DE PEIXES RECIFAIS EM COSTÃO ROCHOSO TROPICAL: ANÁLISE DE SENSIBILIDADE. - 2019. Monografia do Trabalho de Conclusão de Curso Orientador: Prof. Dr. Rafael da Rocha Fortes 1. Territorialidade 2. Ambientes recifais 3. Comportamento agonístico I. Universidade Federal do Estado do Rio de Janeiro II. Comportamento Territorialista do Stegastes fuscus (Cuvier, 1830) e Riqueza de Peixes Recifais em Costão Rochoso Tropical: Análise de Sensibilidade 4 Victor Bastos Teixeira Lupinacci COMPORTAMENTO TERRITORIALISTA DO Stegastes fuscus (Cuvier, 1830) E A ASSEMBLEIA DE PEIXES RECIFAIS EM COSTÃO ROCHOSO TROPICAL: ANÁLISE DE SENSIBILIDADE - 2019. Monografia do Trabalho de Conclusão de Curso apresentada ao Instituto de Biociências da Universidade Federal do Estado do Rio de Janeiro, como parte dos requisitos à obtenção do título de Bacharel em Ciências Ambientais. Aprovada em ___ de _____________ de ______. ____________________________________________________ Prof. Dr. Rafael da Rocha Fortes (Orientador) Universidade Federal do Estado do Rio de Janeiro - UNIRIO ____________________________________________________ Dr. -
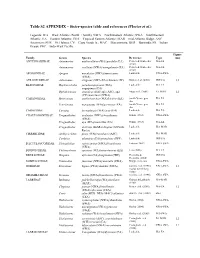
Table S2 APPENDIX – Sister-Species Table and References (Floeter Et Al.)
Table S2 APPENDIX – Sister-species table and references (Floeter et al.) Legends: WA = West Atlantic (North + South); NWA = Northwestern Atlantic; SWA = Southwestern Atlantic; EA = Eastern Atlantic; TEA = Tropical Eastern Atlantic; MAR = mid-Atlantic Ridge; ASC = Ascension; SHE = St. Helena; CV = Cape Verde Is.; MAC = Macaronesia; BER = Bermuda; IO = Indian Ocean; IWP = Indo-West Pacific. Figure Family Genus Species Reference Type (ms) ANTENNARIIDAE Antennarius multiocellatus (WA)/pardalis (EA) Pietsch & Grobecker WA-EA (1987) Antennarius ocellatus (NWA)/senegalensis (EA) Pietsch & Grobecker WA-EA (1987) APOGONIDAE Apogon maculatus (NWA)/americanus Look alike NWA-SWA (SWA) AULOSTOMIDAE Aulostomus strigosus (SWA-EA)/chinensis (IP) Bowen et al. (2001) IWP-EA 12 BLENNIIDAE Hypleurochilus pseudoaequipinnis (WA)/ Look alike WA-EA aequipinnis (EA) Ophioblennius atlanticus (EA)/ sp1 (ASC), sp2 Muss et al. (2001) EA-MAR 12 (CV)/macclurei (NWA) CARANGIDAE Hemicaranx amblyrhynchus (WA)/bicolor (EA) Smith-Vaniz, pers WA-EA obs. Trachinotus marginatus (WA)/goreensis (EA) Smith-Vaniz, pers WA-EA obs. CARAPIDAE Carapus bermudensis (WA)/acus (EA) Look alike WA-EA CHAETODONTIDAE Prognathodes aculeatus (NWA)/brasiliensis Hubbs (1963) NWA-SWA (SWA) Prognathodes aya (WA)/marcellae (EA) Hubbs (1963) WA-EA Prognathodes dichrous (MAR)/obliquus (St Paul's Look alike WA-MAR Rocks) CIRRHITIDAE Amblycirrhitus pinos (WA)/earnshawi (ASC) Look alike WA-MAR Cirrhitus atlanticus (EA)/pinnulatus (IWP) Look alike IWP-EA DACTYLOSCOPIDAE Platygillellus rubrocinctus (NWA)/brasiliensis Feitoza (2002) NWA-SWA (SWA) DIODONTIDAE Chilomycterus spinosus (WA)/mauretanicus (EA) Leis (2006) WA-EA DREPANIDAE Drepane africana (EA)/longimana (IWP) Heemstra & IWP-EA Heemstra (2004) GOBIESOCIDAE Tomicodon fasciatus (NWA)/australis (SWA) Briggs, pers com NWA-SWA GOBIIDAE Elacatinus figaro (SWA)/randalli (NWA) Sazima et al. -

Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes
Old Dominion University ODU Digital Commons Biological Sciences Theses & Dissertations Biological Sciences Summer 2016 Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes Christi Linardich Old Dominion University, [email protected] Follow this and additional works at: https://digitalcommons.odu.edu/biology_etds Part of the Biodiversity Commons, Biology Commons, Environmental Health and Protection Commons, and the Marine Biology Commons Recommended Citation Linardich, Christi. "Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes" (2016). Master of Science (MS), Thesis, Biological Sciences, Old Dominion University, DOI: 10.25777/hydh-jp82 https://digitalcommons.odu.edu/biology_etds/13 This Thesis is brought to you for free and open access by the Biological Sciences at ODU Digital Commons. It has been accepted for inclusion in Biological Sciences Theses & Dissertations by an authorized administrator of ODU Digital Commons. For more information, please contact [email protected]. HOTSPOTS, EXTINCTION RISK AND CONSERVATION PRIORITIES OF GREATER CARIBBEAN AND GULF OF MEXICO MARINE BONY SHOREFISHES by Christi Linardich B.A. December 2006, Florida Gulf Coast University A Thesis Submitted to the Faculty of Old Dominion University in Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE BIOLOGY OLD DOMINION UNIVERSITY August 2016 Approved by: Kent E. Carpenter (Advisor) Beth Polidoro (Member) Holly Gaff (Member) ABSTRACT HOTSPOTS, EXTINCTION RISK AND CONSERVATION PRIORITIES OF GREATER CARIBBEAN AND GULF OF MEXICO MARINE BONY SHOREFISHES Christi Linardich Old Dominion University, 2016 Advisor: Dr. Kent E. Carpenter Understanding the status of species is important for allocation of resources to redress biodiversity loss. -

Deep Reefs Are Not Refugium for Shallow‐Water
Received: 9 November 2020 | Revised: 28 January 2021 | Accepted: 3 February 2021 DOI: 10.1002/ece3.7336 ORIGINAL RESEARCH Deep reefs are not refugium for shallow- water fish communities in the southwestern Atlantic Aline P. M. Medeiros1 | Beatrice P. Ferreira2 | Fredy Alvarado3 | Ricardo Betancur- R4,5 | Marcelo O. Soares6,7,8 | Bráulio A. Santos9 1Programa de Pós- Graduação em Ciências Biológicas, Universidade Federal da Paraíba, João Pessoa, PB, Brazil 2Departamento de Oceanografia, Centro de Tecnologia, Universidade Federal de Pernambuco, Recife, Brazil 3Departamento de Agricultura, Centro de Ciências Humanas, Sociais e Agrárias, Universidade Federal da Paraíba, Bananeiras, PB, Brazil 4Department of Biology, The University of Oklahoma, Norman, OK, USA 5Department of Vertebrate Zoology, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA 6Instituto de Ciências do Mar- LABOMAR, Universidade Federal do Ceará, Meireles, Brazil 7Institut de Ciència i Tecnologia Ambientals (ICTA), Universitat Autònoma de Barcelona (UAB), Barcelona, Spain 8Dipartimento di Scienze e Tecnologie Biologiche e Ambientali (DISTEBA), Università del Salento, Lecce, Italy 9Departamento de Sistemática e Ecologia, Centro de Ciências Exatas e da Natureza, Universidade Federal da Paraíba, João Pessoa, PB, Brazil Correspondence Bráulio A. Santos, Departamento de Abstract Sistemática e Ecologia, Centro de Ciências 1. The deep reef refugia hypothesis (DRRH) predicts that deep reef ecosystems may Exatas e da Natureza, Universidade Federal da Paraíba, -

Universidade Federal De Santa Catarina Centro De Ciências Biológicas Programa De Pós-Graduação Em Ecologia
UNIVERSIDADE FEDERAL DE SANTA CATARINA CENTRO DE CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA ANDREA DALBEN SOARES ESTRUTURA DE COMUNIDADE E DISTRIBUIÇÃO VERTICAL DE PEIXES CRIPTOBÊNTICOS EM ILHAS COSTEIRAS DE SANTA CATARINA, SUL DO BRASIL Florianópolis/SC 2010 ANDREA DALBEN SOARES ESTRUTURA DE COMUNIDADE E DISTRIBUIÇÃO VERTICAL DE PEIXES CRIPTOBÊNTICOS EM ILHAS COSTEIRAS DE SANTA CATARINA, SUL DO BRASIL Dissertação apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal de Santa Catarina, para a obtenção do título de Mestre em Ecologia. Área de concentração : Ecologia, Bases Ecológicas para o Manejo e Conservação de Ecossistemas Costeiros. Orientador: Prof. Dr. Sergio R. Floeter Florianópolis/SC 2010 AGRADECIMENTOS À minha mãe e ao meu pai (in memorian) por me incentivarem e proporcionarem meios para que eu estudasse “aquilo que eu gostasse”, nunca pressionando para que essa escolha fosse direcionada a uma carreira “com um maior mercado de trabalho”, permitindo assim que hoje eu possa ter concluído um trabalho relacionado à ecologia de uns peixinhos pequenininhos que quase ninguém conhece! A Sérgio R. Floeter, por ter acreditado e confiado em mim desde o princípio de um projeto que mais parecia de doutorado (quem sabe agora!?). Agradeço também pela FORÇA nas correções e compreensão durante minhas viagens. Espero ter correspondido. Ao pessoal de campo que me ajudou muito mesmo quando a água estava 14°C: Anderson Batista, Ana Liedke, Igor Pinheiro, Fabrício Richmond. Aos membros da banca (Sonia Buck, Carlos Rangel e Barbara Segal) pelas sugestões e melhorias para este trabalho. À Cecilia Kotzias pelas correções do inglês, ao pessoal do LBMM (www.lbmm.ufsc.br) por todas as sugestões.