Small Freshwater Thalassiosiroid Diatoms From
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CNC/IUGG: 2019 Quadrennial Report
CNC/IUGG: 2019 Quadrennial Report Geodesy and Geophysics in Canada 2015-2019 Quadrennial Report of the Canadian National Committee for the International Union of Geodesy and Geophysics Prepared on the Occasion of the 27th General Assembly of the IUGG Montreal, Canada July 2019 INTRODUCTION This report summarizes the research carried out in Canada in the fields of geodesy and geophysics during the quadrennial 2015-2019. It was prepared under the direction of the Canadian National Committee for the International Union of Geodesy and Geophysics (CNC/IUGG). The CNC/IUGG is administered by the Canadian Geophysical Union, in consultation with the Canadian Meteorological and Oceanographic Society and other Canadian scientific organizations, including the Canadian Association of Physicists, the Geological Association of Canada, and the Canadian Institute of Geomatics. The IUGG adhering organization for Canada is the National Research Council of Canada. Among other duties, the CNC/IUGG is responsible for: • collecting and reconciling the many views of the constituent Canadian scientific community on relevant issues • identifying, representing, and promoting the capabilities and distinctive competence of the community on the international stage • enhancing the depth and breadth of the participation of the community in the activities and events of the IUGG and related organizations • establishing the mechanisms for communicating to the community the views of the IUGG and information about the activities of the IUGG. The aim of this report is to communicate to both the Canadian and international scientific communities the research areas and research progress that has been achieved in geodesy and geophysics over the last four years. The main body of this report is divided into eight sections: one for each of the eight major scientific disciplines as represented by the eight sister societies of the IUGG. -

Pingualuit Crater Lake): Diatom Inferred Paleoenvironmental Record for Three Previous Interglacial Periods
THE “CRYSTAL EYE OF NUNAVIK” (PINGUALUIT CRATER LAKE): DIATOM INFERRED PALEOENVIRONMENTAL RECORD FOR THREE PREVIOUS INTERGLACIAL PERIODS 1,2Jessica L. Black, 1Sonja Hausmann, 2Reinhard Pienitz, 3Veli-Pekka Salonen, 4Guillame St-Onge, 5Michel Bouchard, 6Laura Cunningham, 7Pierre Francus, and 8Michel Lamothe 1Department of Geosciences, University of Arkansas, 113 Ozark Hall, Fayetteville, AR 72701, [email protected], 2Université Laval, Québec, G1V 0A6, Canada, 3Department of Geology, Univ of Helsinki, P.O. BOX 64, Helsinki, FIN-00014, Finland, 4ISMER/GEOTOP, University of Quebec at Rimouski, Rimouski, QC G5L 3A1, Canada, 5Centre des Technologies de l'Environment de Tunis (CITET), Université de Montréal, Montréal, QC H3T 1J4, Canada, 6Abisko Scientific Research Station, Umea University, Abisko, SE-981 07, Sweden, 7Centre Eau, Terre et Environnement, Institut national de la recherche scientifique, 490 rue de la couronne, Québec, QC G1K 9A9, Canada 8Département des sciences de la Terre et de l’Atmosphère, Université du Québec à Montréal, CP 8888 Centre-ville, Montréal, QC H3C 3P8, Canada The sediments of the 1.4. Ma old Pingualuit Crater Lake known as the “Crystal Eye of Nunavik” offer the unique opportunity to study terrestrial climate dynamics not only during the postglacial period, but over several hundreds of thousands of years as its deep sediment infill yields an uninterrupted arctic paleoclimate record. The Pingualuit meteoritic crater (Nunavik, Canada; 61o17' N, 73o41' W) is located in the northernmost part of the Ungava Peninsula in northern Quebec - close to the area where the Laurentide Ice Sheet reached maximum thickness during the last (Wisconsinan) glaciation. In May, 2007 ~10 m of sediments was recovered from the crater lake at a water depth of 270 m using a UWITEC piston percussion corer system. -

Deglacial and Postglacial Evolution of the Pingualuit Crater Lake Basin, Northern Québec (Canada)
Geomorphology 248 (2015) 327–343 Contents lists available at ScienceDirect Geomorphology journal homepage: www.elsevier.com/locate/geomorph Deglacial and postglacial evolution of the Pingualuit Crater Lake basin, northern Québec (Canada) Pierre-Arnaud Desiage a,b,⁎, Patrick Lajeunesse b,c, Guillaume St-Onge a,b, Alexandre Normandeau b, Grégoire Ledoux c,HervéGuyarda,b,d, Reinhard Pienitz c a Institut des sciences de la mer de Rimouski (ISMER), Canada Research Chair in Marine Geology, Université du Québec à Rimouski, Rimouski, Canada b GEOTOP Research Center, Canada c Centre d'études Nordiques (CEN) & Département de Géographie, Université Laval, Québec, Canada d Institut de Physique du Globe de Paris, Sorbonne Paris Cité, Université Paris Diderot, UMR CNRS 7154, Paris, France article info abstract Article history: The Pingualuit Crater, located in the Ungava Peninsula (northern Québec, Canada) is a 1.4-Ma-old impact crater Received 15 January 2015 hosting a ~245-m-deep lake. The lake has a great potential to preserve unique paleoclimatic and paleoecological Received in revised form 9 July 2015 sedimentary records of the last glacial/interglacial cycles in the terrestrial Canadian Arctic. In order to investigate Accepted 10 July 2015 the stratigraphy in the lake and the late Quaternary glacial history of the Pingualuit Crater, this study compiles Available online 29 July 2015 data from three expeditions carried out in May 2007 (~9-m-long sediment core), in August 2010 (~50 km of seismic lines), and in September 2012 (high-resolution terrestrial LiDAR topography of the inner slopes). Despite Keywords: fi lake levels the weak penetration (~10 m) of the 3.5-kHz subbottom pro ling caused by the presence of boulders in the Mass movements sedimentary column, seismic data coupled with the stratigraphy established from the sediment core enabled Laurentide Ice Sheet the identification of two glaciolacustrine units deposited during the final stages of the Laurentide Ice Sheet Sedimentological processes (LIS) retreat in the crater. -

Results of Recent Sediment Drilling Activities in Deep Crater Lakes
of the subtropical region of South Ameri- beyond the instrumental record, as well del Puerto, L, García-Rodríguez, F., Inda H., Bracco, R., Castiñeira, C., J.B. and Adams, J.B., 2006: Paleolimnological evidence of Holocene ca (i.e., the transition between the tropics as to design new strategies for the sus- paleoclimatic changes in Lake Blanca, southern Uruguay, Journal and the extra tropics) in deciphering the tainable development of ecosystems and of Paleolimnology, 36: 151-163. forcing factors of past changes in atmo- natural resources. García-Rodríguez, F., 2006: Inferring paleosalinity changes using the chrysophyte cyst to diatom ratio in coastal shallow temperate/ spheric circulation. Both Argentinean and subtropical lagoons influenced by sea level changes, Journal of Uruguayan Holocene paleolimnologi- Acknowledgements Paleolimnology, 36,165-173. PEDECIBA, SNI-ANII and CSIC-Program Piovano, E., Ariztegui, D., Bernasconi, S.M. and McKenzie, J.A., 2004: cal records indicate similar paleoclimatic Stable isotopic record of hydrological changes in subtropical trends, i.e. dominant dry conditions were “Contratación de investigadores provenientes Laguna Mar Chiquita (Argentina) over the last 230 years, The del exterior” (Uruguay) are thanked for finan- observed during cold phases, whereas Holocene, 14(4): 525-535. cial support. Part of this study was supported Piovano, E.L., Ariztegui, D., Córdoba, F., Cioccale, M. and Sylvestre, F., wet conditions prevailed during warm by Argentina through funding of CONICET (PIP 2009: Hydrological variability in South America below the Tropic climatic phases. To our knowledge, this is 5947; PIP 112-200801-00808), FONCYT (PICT Nº of Capricorn (Pampas and Patagonia, Argentina) during the last 13.0 In: Vimeux, F. -

Uncorrected Proof
Quaternary Science Reviews xxx (2018) xxx-xxx Contents lists available at ScienceDirect Quaternary Science Reviews journal homepage: www.elsevier.com Late-Quaternary glacial to postglacial sedimentation in three adjacent fjord-lakes of the Québec North Shore (eastern Canadian Shield) Antoine G. Poiréa, ∗, Patrick Lajeunessea, Alexandre Normandeaua, b, Pierre Francusc, Guillaume St-Onged, Obinna P. Nzekwec a Département de géographie & Centre d’études nordiques, Université Laval, Québec, Qc G1V 0A6, Canada b Geological Survey of Canada (Atlantic), Bedford Institute of Oceanography, P.O. Box 1006, Dartmouth, Nova Scotia B2Y 4A2, Canada c Centre Eau Terre Environnement, Institut National de la Recherche Scientifique, Québec, Qc G1K 9A9, Canada, Canada Research Chair in Environmental Sedimentology & GEOTOP d Institut des sciences de la mer de Rimouski, Université du Québec à Rimouski, Rimouski, Qc, Canada, Canada Research Chair in MarinePROOFGeology & GEOTOP ARTICLE INFO ABSTRACT Article history: High-resolution swath bathymetry imagery allowed mapping in great detail the sublacustrine geomorphology Received 23 June 2017 of lakes Pentecôte, Walker and Pasteur, three deep adjacent fjord-lakes of the Québec North Shore (eastern Received in revised form 31 January 2018 Canada). These sedimentary basins have been glacio-isostatically uplifted to form deep steep-sided elongated Accepted 11 February 2018 lakes. Their key geographical position and limnogeological characteristics typical of fjords suggest excep- Available online xxx tional potential -

Project Caribou an Educator’S Guide to Wild Caribou of North America
An Educator’s Guide to Wild Caribou of North America Copyright 2018 This book may be reproduced for educational purposes only. The illustrations may be duplicated solely for instructional use of this material. Reproduction for any other purpose is strictly prohibited without written permission from the publisher. For more information please contact: Environmental Education and Youth Programs Government of Yukon, Department of Environment Conservation Officer Services Box 2703, Whitehorse, Yukon Y1A 2C6 (867) 667-3675 or 1-800-661-0408 fax (867) 393-6206 [email protected] ISBN: 978-1-55362-794-4 Photo and illustration credits Government of Yukon, Department of Environment Government of Northwest Territories 1, 4, 6, 7, 8, 11, 12, 13, 16, 17, 19, 23, 25, 26, 27, 29, 31, 55, 65, 78, 87, 89 90, 94, 98, 99, 100, 102, 122 Government of Nunavut Doug Urquhart (illustrations) 132 3, 20, 26, 27, 29, 31, 34, 36, 42, 53, 63, 68, 75, 84, 98, 121 Joyce Majiski Government of Canada, Canadian Wildlife Service Cover, i 2, 9, 15, 18, 22, 24 Tanya Handley (illustrations) vii, 37, 40, 43, 102, 103 Jennifer Staniforth (illustrations) 4, 5, 10, 63 Beverly and Qamanirjuaq Caribou Management Board (illustrations) 47, 48, 49, 50 Northern Contaminants Committee 37, 40, 42, 43 Government of Quebec 112, 115 Project Caribou An Educator’s Guide to Wild Caribou of North America Project manager: Remy Rodden, Government of Yukon, Department of Environment Consultants: Darielle Talarico, Arctic Vision, Whitehorse, Yukon Principal writer: Kirsten Madsen, Whitehorse, Yukon Original illustrations: Jennifer Staniforth, Doug Urquhart, Tanya Handley Cover: Joyce Majiski Editing and production: Walker LeBrun Creative Services, Whitehorse, Yukon Government of Yukon, Department of Environment (2018 edition) Beverly and Qamanirjuaq Caribou Management Board i Project Caribou Acknowledgments Project Caribou is the result of the Sharon Russell Nova Scotia efforts of many individuals. -

Arctic Workshop 2013 Program Summary
Arctic Workshop 2013 Program Summary SUNDAY 10 MARCH 9:00 – 4:00 Pre-workshop field trip Hotel Lobby 7:00 – 9:30 Evening reception, Check-In, Level 11 Registration (load presentations onto computer) MONDAY 11 MARCH 8:00 – 8:30 Check-in & Registration Level 2 Reading Room (load presentations onto computer) 8:45 Welcome & Introduction Level 2 Reading Room 9:00 Paleoceanography I Level 2 Reading Room 10:10 Morning Break Level 1 Auditorium 10:40 Paleoceanography II Level 2 Reading Room 12:00 LUNCH - 1:00 pm Glacial History Level 2 Reading Room 3:50 Afternoon Break Level 1 Auditorium 4:00 POSTERS I (odd-numbered posters) Level 1 Auditorium 6:00 End of session Level 1 Auditorium 6:30 Non-student Dinner Julie B-G’s house 7:00 Student-only Pizza Party University Club TUESDAY 12 MARCH 9:00 Paleolimnology Level 2 Reading Room 10:30 Morning Break Level 1 Auditorium 10:50 Modelling Level 2 Reading Room 12:10 LUNCH - 1:20 pm Special Talk (Kevin Knobloch, UCS) Level 2 Reading Room 1:50 Glaciology Level 2 Reading Room 3:45 Afternoon Break Level 1 Auditorium 4:00 Chasing Ice – documentary film ISB Auditorium 5:15 Afternoon Break Level 1 Auditorium 5:30 POSTERS II (even numbered posters) Level 1 Auditorium 7:00 End of session Level 1 Auditorium 7:00 Workshop Banquet Level 1 Auditorium Keynote: John H. England Level 1 Auditorium WEDNESDAY 13 MARCH 8:45 Paleoclimatology I Level 2 Reading Room 10:35 Morning Break Level 1 Auditorium 10:50 Paleoclimatology II Level 2 Reading Room 12:40 End of Meeting. -

Maan Kosmiset Arvet - Törmäysrakenteet
Hinta - Pris 5 € ASTERISKI 2014 Tähtitieteellinen yhdistys Vaasan Andromeda ry | Astronomiska föreningen Vasa Andromeda rf Jäsenlehti - Medlemstidning Satu Hietala, Geologian tutkimuskeskus MAAN KOSMISET ARVET - TÖRMÄYSRAKENTEET Meteoriihen viisivuotis Internationell ring av Styrning av sonder i syntymäpäiviä vietettiin forskare vill borra i rymden kesäkuun 29 päivänä. Söderfjärden Asteriski 2014 1 Meteoriihen viisivuotis syntymäpäiviä vietettiin lauantaina kesäkuun 29 päivänä. Juhlapuhujana Esko Valtaoja Kuva Kaj Höglund Kuva: Miikis RC METEORIIHI 5vuotta Kuva Kaj Höglund 4 3 Terveiset puheenjohtajalta 4 Meteoriihi 5 vuotta 6 Maan kosmiset arvet - törmäysrakenteet 14 Styrning av sonder i rymden 14 Nova Delphini 2013 16 Internationell ring av forskare vill borra i Söderfjärden 17 Kansainvälinen tutkijaryhmä haluaa kairata Söderfjärdenillä 18 Hei, tämä on minun tarinani 6 19 Supernovat SN 2013ej M74- ja SN2014J M81-galaksissa MAAN KOSMISET ARVET - 20 Nyinköpt utrustning 2013 | Uusia varustehankintoja 2013 TÖRMÄYSRAKENTEET 20 Andromeda vierailulla Lakeuden ursan tornilla Kuvassa uusi lohkarelöytö 20 Leader-hanke vuodelta 2013 Söderfjärdeniltä, 21 Vad är Celestia? joka on mahdollisesti sueviittia eli törmäyksessä ilmaan lentä- 22 Ett ”vanvettsföretag” nyttä kiviainesta. 23 Liity Andromedan jäseneksi / Bli medlem i Andromeda / sisältö 2014 innehåll Apply for membership Hinta - Pris 5 € ASTERISKI 2014 Tähtitieteellinen yhdistys Vaasan Andromeda ry | Astronomiska föreningen Vasa Andromeda rf Jäsenlehti - Medlemstidning JULKAISIJA Tähtitieteellinen -

Guyard-ET-AL JQSR Proof.Pdf
Quaternary Science Reviews xxx (2011) 1e16 Contents lists available at SciVerse ScienceDirect Quaternary Science Reviews journal homepage: www.elsevier.com/locate/quascirev New insights into Late Pleistocene glacial and postglacial history of northernmost Ungava (Canada) from Pingualuit Crater Lake sediments Hervé Guyard a,b,*, Guillaume St-Onge a,b, Reinhard Pienitz c, Pierre Francus b,d, Bernd Zolitschka e, Garry K.C. Clarke f, Sonja Hausmann g, Veli-Pekka Salonen h, Patrick Lajeunesse c, Grégoire Ledoux c, Michel Lamothe i a Canada Research Chair in Marine Geology, Institut des sciences de la mer de Rimouski (ISMER), UQAR, Rimouski, Canada b GEOTOP Research Center, Canada c Centre d’études nordiques (CEN) & Département de Géographie, Université Laval, Québec, Canada d Institut National de la Recherche Scientifique e Centre Eau, Terre, Environnement, Québec, Canada e Institut für Geographie, University of Bremen, Germany f Department of Earth and Ocean Sciences, University of British Columbia, Canada g Department of Geosciences, University of Arkansas, USA h Department of Geosciences and Geography, University of Helsinki, Finland i Département des Sciences de la Terre et de l’Atmosphère, UQAM, Montréal, Canada article info abstract Article history: The Pingualuit Crater was formed by a meteoritic impact ca. 1.4 million years ago in northernmost Received 8 April 2011 Ungava (Canada). Due to its geographical position near the center of successive North American ice Received in revised form sheets and its favorable morphometry, the Pingualuit Crater Lake (water depth ¼ 246 m) promises to 6 October 2011 yield a unique continuous sedimentary sequence covering several glacial/interglacial cycles in the Accepted 7 October 2011 terrestrial Canadian Arctic. -
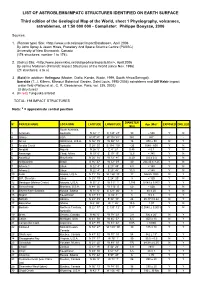
LIST of ASTROBLEMS/IMPATC STRUCTURES IDENTIFIED on EARTH SURFACE Third Edition of the Geological Map of the World, Sheet 1 Physi
LIST OF ASTROBLEMS/IMPATC STRUCTURES IDENTIFIED ON EARTH SURFACE Third edition of the Geological Map of the World, sheet 1 Physiography, volcanoes, astroblemes, at 1:50 000 000 - Compilator: Philippe Bouysse, 2006 Sources: 1. (Roman type) Site: <http://www.unb.ca/passc/ImpactDatabase>, April 2006 By John Spray & Jason Hines, Planetary And Space Science Centre (PASSC) University of New Brunswick, Canada (174 structures: number 1 to 174), 2. (Italics) Site: <http://www.somerikko.net/old/geo/imp/impacts.htm>, April 2006 By Jarmo Moilanen (Finland): Impact Structures of the World (since Nov. 1996) (21 structures: a to u) 3. (Bold) In addition: Velingara (Master, Diallo, Kande, Wade, 1999, South Africa/Senegal), Iturralde (T. J. Killeen, Missouri Botanical Garden, Saint Louis, 1995-2006) astroblems and Gilf Kebir impact crater field (Paillou et al., C. R. Géoscience, Paris, vol. 335, 2003). (3 structures) 4. (In red) Tunguska airblast TOTAL: 198 IMPACT STRUCTURES Nota: * = approximate central position DIAMETER N° CRATER NAME LOCATION LATITUDE LONGITUDE Age (Ma)* EXPOSED DRILLED (km) South Australia, 1 Acraman Australia S 32° 1' E 135° 27' 90 ~ 590 Y N a Alamo USA N 37°30' W 116°30' 190 367 - - 2 Ames Oklahoma, U.S.A. N 36° 15' W 98° 12' 16 470 ± 30 N Y 3 Amelia Creek Australia S 20° 55' E 134 ° 50' ~20 1640 - 600 Y N 4 Amguid Algeria N 26° 5' E 4° 23' 0.45 < 0.1 Y N 5 Aorounga Chad, Africa N 19° 6' E 19° 15' 12.6 < 345 Y N 6 Aouelloul Mauritania N 20° 15' W 12° 41' 0.39 3.0 ± 0.3 Y N 7 Araguainha Brazil S 16° 47' W 52° 59' 40 244.40 ± 3.25 Y N 8 Arkenu 1 Libya N 22° 4' E 23° 45' 6.8 < 140 Y N 9 Arkenu 2 Libya N 22° 4' E 23° 45' 10,3 < 140 Y N 10 Avak Alaska, U.S.A. -

Québec Aventure Plein
QUÉBECoutdoors North America / Canada / Québec Nature in Québec beckons, bewitches and astounds. And our wildlife couldn’t be more diverse! 2 3 1 4 CARIBOU FISHING OUTFITTERS REFUGE TIPI INUKSHUK NATURE CULTURE KAYAKING HIKING BELUGA ADVENTURE HUSKY SNOWMOBILING FOREST CAMPING INUIT TOUNDRA INN AUTHENTICITY 1 3. Parcs Québec (National Parks) 4. Northern Québec Abitibi-Témiscamingue / Eeyou Istchee – Baie-James / Nunavik 7. Aventure Écotourisme Québec 8. Western Québec Outaouais / Laurentides / Montréal 10. Mauricie / Lanaudière 12. Québec Outfitters Federation 13. Québec City & Area / Charlevoix 16. Québec Aboriginal Tourism 17. Saguenay / Lac-Saint-Jean 21. Maritime Québec Gaspésie / Lower St. Lawrence / Noth Shore 2 THE NATIONAL PARKS OF THE PARCS QUÉBEC NETWORK 24 25 1 Parc national d’Aiguebelle 9 Parc national de l’Île-Bonaventure- 17 Parc national du Mont-Tremblant 2 Parc national d’Anticosti et-du-Rocher-Percé 18 Parc national des Monts-Valin 3 Parc national du Bic 10 Parc national des Îles-de-Boucherville 19 Parc national d’Oka 4 Parc national du Fjord-du-Saguenay 11 Parc national de la Jacques-Cartier 20 Parc national de Plaisance 5 Parc national de Frontenac 12 Parc national du Lac-Témiscouata 21 Parc national de la Pointe-Taillon 6 Parc national de la Gaspésie 13 Parc national de Miguasha 22 Parc national de la Yamaska 7 Parc national des Grands-Jardins 14 Parc national du Mont-Mégantic 23 Parc marin du Saguenay–Saint-Laurent 8 Parc national des Hautes-Gorges- 15 Parc national du Mont-Orford 24 Parc national des Pingualuit de-la-Rivière-Malbaie 16 Parc national du Mont-Saint-Bruno 25 Parc national Kuururjuaq QUÉBEC NATIONAL PARKS Why visit Québec’s National Parks? Simple. -

Évolution Tardi-Quaternaire Et Morpho-Stratigraphie Des
UNIVERSITÉ DU QUÉBEC À RIMOUSKI Évolution tardi-quaternaire et morpho-stratigraphie des sédiments du cratère des Pingualuit : variation du niveau du lac, origine des mouvements de masse et influence de la glaciation WlsconSlnlenne. .. Mémoire présenté dans le cadre du programme de maîtrise en océanographie en vue de l'obtention du grade de maître ès sciences PAR © PIERRE-ARNAUD DESIAGE Janvier 2014 UNIVERSITÉ DU QUÉBEC À RlMOUSKI Service de la bibliothèque Avertissement La diffusion de ce mémoire ou de cette thèse se fait dans le respect des droits de son auteur, qui a signé le formulaire « Autorisation de reproduire et de diffuser un rapport, un mémoire ou une thèse». En signant ce formulaire, l'auteur concède à l'Université du Québec à Rimouski un e licence non exclusive d' utili sation et de publication de la totalité ou d' un e partie importante de so n travail de recherche pour des fins pédagogiques et non com merciale s. Plus précisément, l'auteur autorise l' Université du Québec à Rimouski à reproduire, diffuser, prêter, distribuer ou vendre des copies de son travail de recherche à des fins non co mmerciales sur quelque support que ce soit, y co mpri s l' Internet. Cette licence et cette autorisation n'entraînent pas une renonciation de la part de l'auteur à ses droits moraux ni à ses droits de propriété intellectuelle. Sauf entente contraire, l'auteur co nserve la liberté de diffuser et de commercialiser ou non ce travail dont il possède un exemplaire. 11 III Composition du jury : Jean Carlos Montero Serrano, président du jury, UQAR-ISMER Guillaume St-Onge, directeur de recherche, UQAR-ISMER Patrick Lajeunesse, codirecteur de recherche, Université Laval Pierre Francus, examinateur externe, INRS Dépôt initial le 28 janvier 2014 Dépôt final le 13 juin 2014 IV vu Prenez le plus rêveur des rêveurs, plantez-le sur ses pattes, faites marcher ses pieds..