4 the Anatomy and Physiology of the Diaphragm
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Part 1 the Thorax ECA1 7/18/06 6:30 PM Page 2 ECA1 7/18/06 6:30 PM Page 3
ECA1 7/18/06 6:30 PM Page 1 Part 1 The Thorax ECA1 7/18/06 6:30 PM Page 2 ECA1 7/18/06 6:30 PM Page 3 Surface anatomy and surface markings The experienced clinician spends much of his working life relating the surface anatomy of his patients to their deep structures (Fig. 1; see also Figs. 11 and 22). The following bony prominences can usually be palpated in the living subject (corresponding vertebral levels are given in brackets): •◊◊superior angle of the scapula (T2); •◊◊upper border of the manubrium sterni, the suprasternal notch (T2/3); •◊◊spine of the scapula (T3); •◊◊sternal angle (of Louis) — the transverse ridge at the manubrio-sternal junction (T4/5); •◊◊inferior angle of scapula (T8); •◊◊xiphisternal joint (T9); •◊◊lowest part of costal margin—10th rib (the subcostal line passes through L3). Note from Fig. 1 that the manubrium corresponds to the 3rd and 4th thoracic vertebrae and overlies the aortic arch, and that the sternum corre- sponds to the 5th to 8th vertebrae and neatly overlies the heart. Since the 1st and 12th ribs are difficult to feel, the ribs should be enu- merated from the 2nd costal cartilage, which articulates with the sternum at the angle of Louis. The spinous processes of all the thoracic vertebrae can be palpated in the midline posteriorly, but it should be remembered that the first spinous process that can be felt is that of C7 (the vertebra prominens). The position of the nipple varies considerably in the female, but in the male it usually lies in the 4th intercostal space about 4in (10cm) from the midline. -

Diaphragm and Intercostal Muscles
Diaphragm and intercostal muscles Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology Skeletal System Adult Human contains 206 Bones 2 parts: Axial skeleton (axis): Skull, Vertebral column, Thoracic cage Appendicular skeleton: Bones of upper limb Bones of lower limb Dr. Heba Kalbouneh Structure of Typical Vertebra Body Vertebral foramen Pedicle Transverse process Spinous process Lamina Dr. Heba Kalbouneh Superior articular process Intervertebral disc Dr. Heba Inferior articular process Dr. Heba Facet joints are between the superior articular process of one vertebra and the inferior articular process of the vertebra directly above it Inferior articular process Superior articular process Dr. Heba Kalbouneh Atypical Vertebrae Atlas (1st cervical vertebra) Axis (2nd cervical vertebra) Dr. Heba Atlas (1st cervical vertebra) Communicates: sup: skull (atlanto-occipital joint) inf: axis (atlanto-axial joint) Atlas (1st cervical vertebra) Characteristics: 1. no body 2. no spinous process 3. ant. & post. arches 4. 2 lateral masses 5. 2 transverse foramina Typical cervical vertebra Specific to the cervical vertebra is the transverse foramen (foramen transversarium). is an opening on each of the transverse processes which gives passage to the vertebral artery Thoracic Cage - Sternum (G, sternon= chest bone) -12 pairs of ribs & costal cartilages -12 thoracic vertebrae Manubrium Body Sternum: Flat bone 3 parts: Xiphoid process Dr. Heba Kalbouneh Dr. Heba Kalbouneh The external intercostal muscle forms the most superficial layer. Its fibers are directed downward and forward from the inferior border of the rib above to the superior border of the rib below The muscle extends forward to the costal cartilage where it is replaced by an aponeurosis, the anterior (external) intercostal membrane Dr. -

Slipping Rib Syndrome
Slipping Rib Syndrome Jackie Dozier, BS Edited by Lisa E McMahon, MD FACS FAAP David M Notrica, MD FACS FAAP Case Presentation AA is a 12 year old female who presented with a 7 month history of right-sided chest/rib pain. She states that the pain was not preceded by trauma and she had never experienced pain like this before. She has been seen in the past by her pediatrician, chiropractor, and sports medicine physician for her pain. In May 2012, she was seen in the ER after having manipulations done on her ribs by a sports medicine physician. Pain at that time was constant throughout the day and kept her from sleeping. However, it was relieved with hydrocodone/acetaminophen in the ER. Case Presentation Over the following months, the pain became progressively worse and then constant. She also developed shortness of breath. She is a swimmer and says she has had difficulty practicing due to the pain and SOB. AA was seen by a pediatric surgeon and scheduled for an interventional pain management service consult for a test injection. Following good temporary relief by local injection, she was scheduled costal cartilage removal to treat her pain. What is Slipping Rib Syndrome? •Slipping Rib Syndrome (SRS) is caused by hypermobility of the anterior ends of the false rib costal cartilages, which leads to slipping of the affected rib under the superior adjacent rib. •SRS an lead to irritation of the intercostal nerve or strain of the muscles surrounding the rib. •SRS is often misdiagnosed and can lead to months or years of unresolved abdominal and/or thoracic pain. -

Structure of the Human Body
STRUCTURE OF THE HUMAN BODY Vertebral Levels 2011 - 2012 Landmarks and internal structures found at various vertebral levels. Vertebral Landmark Internal Significance Level • Bifurcation of common carotid artery. C3 Hyoid bone Superior border of thyroid C4 cartilage • Larynx ends; trachea begins • Pharynx ends; esophagus begins • Inferior thyroid A crosses posterior to carotid sheath. • Middle cervical sympathetic ganglion C6 Cricoid cartilage behind inf. thyroid a. • Inferior laryngeal nerve enters the larynx. • Vertebral a. enters the transverse. Foramen of C 6. • Thoracic duct reaches its greatest height C7 Vertebra prominens • Isthmus of thyroid gland Sternoclavicular joint (it is a • Highest point of apex of lung. T1 finger's breadth below the bismuth of the thyroid gland T1-2 Superior angle of the scapula T2 Jugular notch T3 Base of spine of scapula • Division between superior and inferior mediastinum • Ascending aorta ends T4 Sternal angle (of Louis) • Arch of aorta begins & ends. • Trachea ends; primary bronchi begin • Heart T5-9 Body of sternum T7 Inferior angle of scapula • Inferior vena cava passes through T8 diaphragm T9 Xiphisternal junction • Costal slips of diaphragm T9-L3 Costal margin • Esophagus through diaphragm T10 • Aorta through diaphragm • Thoracic duct through diaphragm T12 • Azygos V. through diaphragm • Pyloris of stomach immediately above and to the right of the midline. • Duodenojejunal flexure to the left of midline and immediately below it Tran pyloric plane: Found at the • Pancreas on a line with it L1 midpoint between the jugular • Origin of Superior Mesenteric artery notch and the pubic symphysis • Hilum of kidneys: left is above and right is below. • Celiac a. -

1 the Thoracic Wall I
AAA_C01 12/13/05 10:29 Page 8 1 The thoracic wall I Thoracic outlet (inlet) First rib Clavicle Suprasternal notch Manubrium 5 Third rib 1 2 Body of sternum Intercostal 4 space Xiphisternum Scalenus anterior Brachial Cervical Costal cartilage plexus rib Costal margin 3 Subclavian 1 Costochondral joint Floating ribs artery 2 Sternocostal joint Fig.1.3 3 Interchondral joint Bilateral cervical ribs. 4 Xiphisternal joint 5 Manubriosternal joint On the right side the brachial plexus (angle of Louis) is shown arching over the rib and stretching its lowest trunk Fig.1.1 The thoracic cage. The outlet (inlet) of the thorax is outlined Transverse process with facet for rib tubercle Demifacet for head of rib Head Neck Costovertebral T5 joint T6 Facet for Tubercle vertebral body Costotransverse joint Sternocostal joint Shaft 6th Angle rib Costochondral Subcostal groove joint Fig.1.2 Fig.1.4 A typical rib Joints of the thoracic cage 8 The thorax The thoracic wall I AAA_C01 12/13/05 10:29 Page 9 The thoracic cage Costal cartilages The thoracic cage is formed by the sternum and costal cartilages These are bars of hyaline cartilage which connect the upper in front, the vertebral column behind and the ribs and intercostal seven ribs directly to the sternum and the 8th, 9th and 10th ribs spaces laterally. to the cartilage immediately above. It is separated from the abdominal cavity by the diaphragm and communicates superiorly with the root of the neck through Joints of the thoracic cage (Figs 1.1 and 1.4) the thoracic inlet (Fig. -

Yagenich L.V., Kirillova I.I., Siritsa Ye.A. Latin and Main Principals Of
Yagenich L.V., Kirillova I.I., Siritsa Ye.A. Latin and main principals of anatomical, pharmaceutical and clinical terminology (Student's book) Simferopol, 2017 Contents No. Topics Page 1. UNIT I. Latin language history. Phonetics. Alphabet. Vowels and consonants classification. Diphthongs. Digraphs. Letter combinations. 4-13 Syllable shortness and longitude. Stress rules. 2. UNIT II. Grammatical noun categories, declension characteristics, noun 14-25 dictionary forms, determination of the noun stems, nominative and genitive cases and their significance in terms formation. I-st noun declension. 3. UNIT III. Adjectives and its grammatical categories. Classes of adjectives. Adjective entries in dictionaries. Adjectives of the I-st group. Gender 26-36 endings, stem-determining. 4. UNIT IV. Adjectives of the 2-nd group. Morphological characteristics of two- and multi-word anatomical terms. Syntax of two- and multi-word 37-49 anatomical terms. Nouns of the 2nd declension 5. UNIT V. General characteristic of the nouns of the 3rd declension. Parisyllabic and imparisyllabic nouns. Types of stems of the nouns of the 50-58 3rd declension and their peculiarities. 3rd declension nouns in combination with agreed and non-agreed attributes 6. UNIT VI. Peculiarities of 3rd declension nouns of masculine, feminine and neuter genders. Muscle names referring to their functions. Exceptions to the 59-71 gender rule of 3rd declension nouns for all three genders 7. UNIT VII. 1st, 2nd and 3rd declension nouns in combination with II class adjectives. Present Participle and its declension. Anatomical terms 72-81 consisting of nouns and participles 8. UNIT VIII. Nouns of the 4th and 5th declensions and their combination with 82-89 adjectives 9. -

Anatomic Connections of the Diaphragm: Influence of Respiration on the Body System
Journal of Multidisciplinary Healthcare Dovepress open access to scientific and medical research Open Access Full Text Article ORIGINAL RESEARCH Anatomic connections of the diaphragm: influence of respiration on the body system Bruno Bordoni1 Abstract: The article explains the scientific reasons for the diaphragm muscle being an important Emiliano Zanier2 crossroads for information involving the entire body. The diaphragm muscle extends from the trigeminal system to the pelvic floor, passing from the thoracic diaphragm to the floor of the 1Rehabilitation Cardiology Institute of Hospitalization and Care with mouth. Like many structures in the human body, the diaphragm muscle has more than one Scientific Address, S Maria Nascente function, and has links throughout the body, and provides the network necessary for breathing. Don Carlo Gnocchi Foundation, 2EdiAcademy, Milano, Italy To assess and treat this muscle effectively, it is necessary to be aware of its anatomic, fascial, and neurologic complexity in the control of breathing. The patient is never a symptom localized, but a system that adapts to a corporeal dysfunction. Keywords: diaphragm, fascia, phrenic nerve, vagus nerve, pelvis Anatomy and anatomic connections The diaphragm is a dome-shaped musculotendinous structure that is very thin (2–4 mm) and concave on its lower side and separates the chest from the abdomen.1 There is a central tendinous portion, ie, the phrenic center, and a peripheral muscular portion originating in the phrenic center itself.2 With regard to anatomic attachments, -

SŁOWNIK ANATOMICZNY (ANGIELSKO–Łacinsłownik Anatomiczny (Angielsko-Łacińsko-Polski)´ SKO–POLSKI)
ANATOMY WORDS (ENGLISH–LATIN–POLISH) SŁOWNIK ANATOMICZNY (ANGIELSKO–ŁACINSłownik anatomiczny (angielsko-łacińsko-polski)´ SKO–POLSKI) English – Je˛zyk angielski Latin – Łacina Polish – Je˛zyk polski Arteries – Te˛tnice accessory obturator artery arteria obturatoria accessoria tętnica zasłonowa dodatkowa acetabular branch ramus acetabularis gałąź panewkowa anterior basal segmental artery arteria segmentalis basalis anterior pulmonis tętnica segmentowa podstawna przednia (dextri et sinistri) płuca (prawego i lewego) anterior cecal artery arteria caecalis anterior tętnica kątnicza przednia anterior cerebral artery arteria cerebri anterior tętnica przednia mózgu anterior choroidal artery arteria choroidea anterior tętnica naczyniówkowa przednia anterior ciliary arteries arteriae ciliares anteriores tętnice rzęskowe przednie anterior circumflex humeral artery arteria circumflexa humeri anterior tętnica okalająca ramię przednia anterior communicating artery arteria communicans anterior tętnica łącząca przednia anterior conjunctival artery arteria conjunctivalis anterior tętnica spojówkowa przednia anterior ethmoidal artery arteria ethmoidalis anterior tętnica sitowa przednia anterior inferior cerebellar artery arteria anterior inferior cerebelli tętnica dolna przednia móżdżku anterior interosseous artery arteria interossea anterior tętnica międzykostna przednia anterior labial branches of deep external rami labiales anteriores arteriae pudendae gałęzie wargowe przednie tętnicy sromowej pudendal artery externae profundae zewnętrznej głębokiej -

Subcostal TAP, Rectus Sheath
Station 4: Traditional Transversus Abdominis Plane (TAP), Subcostal TAP, Rectus Sheath Station Faculty: Traditional TAP: Adam Amundson, MD David Olsen, MD Indications: o Surgeries that involve the lower abdominal segment (below umbilicus) ie: Pfannenstiel incision for cesarean-section. Ultrasound settings and Patient Position o High Frequency linear probe; Low Frequency linear probe if morbidly obese (38-50 mm footprint) o Supine, arms to sides, performed with patient either sedated or under general anesthesia Surface Anatomy landmarks o Rib cage, costal margin o Iliac Crest, pelvic brim o Mid-axillary line Sonoanatomy – o Start by placing the ultrasound probe between the iliac crest and costal margin at the mid-axillary line o Identify the 3 muscular layers (external oblique EO, internal oblique IO, transversus abdominis TA) and the peritoneum o TIPS: EO is generally hyperechoic, IO is the largest, TA is thin, Bowel/Peritoneum may move EO EO IO IO Superior Superior TA TA Bowel Bowel Suggested Injection Technique- TA o In-plane short axis approach, sonographically guide a 21 g 4 inch needle in between the fascial plane of the IO and TA in the posterior corner (see white arrow below). o Inject 20 mL of dilute long acting local anesthetic per side, or 30 mL if unilateral EO EO IO IO Superior Superior TA Bowel Bowel Bowel o TIP: When entering fascial planes between IO and TA the needle tip is often positioned deep and you may need to slowly pull the needle back while giving small aliquots of fluid to help define the spread of the fascial planes. -

Muscles Involved in Respiration
Prof. Ahmed Fathalla Ibrahim Professor of Anatomy College of Medicine King Saud University E-mail: [email protected] OBJECTIVES At the end of the lecture, students should: ▪ Describe the components of the thoracic cage and their articulations. ▪ Describe in brief the respiratory movements. ▪ List the muscles involved in inspiration and in expiration. ▪ Describe the attachments of each muscle to the thoracic cage and its nerve supply. ▪ Describe the origin, insertion, nerve supply of diaphragm. THORACIC CAGE Vertebra Rib THORACIC CAGE ❑Conical in shape ❑Has 2 apertures (openings): 1. Superior (thoracic outlet): narrow, open, continuous with neck 2. Inferior: wide, closed by diaphragm ❑ Formed of: 1. Sternum & costal cartilages: anteriorly 2. Twelve pairs of ribs: laterally 3. Twelve thoracic vertebrae: posteriorly ARTICULATIONS Costovertebral Manubriosternal Intervertebral disc Costochondral Sternocostal Xiphisternal ARTICULATIONS Costovertebral Sternocostal Costochondral Interchondral ARTICULATIONS • Secondary cartilaginous: Manubriosternal joint, Xiphisternal joint and Intervertebral discs. • Primary cartilaginous: 1st Sternocostal joint, Costochondral joints and Interchondral joints. • Plane synovial joints: Costovertebral joints and the rest of Sternocostal joints. RESPIRATORY MOVEMENTS A- MOVEMENTS OF DIAPHRAGM Inspiration Contraction (descent) of diaphragm Increase of vertical diameter of thoracic cavity Relaxation (ascent) of diaphragm) Expiration RESPIRATORY MOVEMENTS B- MOVEMENTS OF RIBS PUMP HANDLE MOVEMENT BUCKET HANDLE -
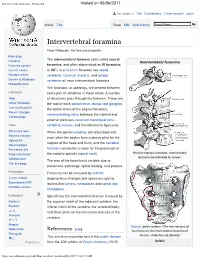
Intervertebral Foramina - Wikipedia Visited on 06/06/2017
Intervertebral foramina - Wikipedia Visited on 06/06/2017 Not logged in Talk Contributions Create account Log in Article Talk Read Edit View history Intervertebral foramina From Wikipedia, the free encyclopedia Main page The intervertebral foramen (also called neural Contents Intervertebral foramina Featured content foramina, and often abbreviated as IV foramina Current events or IVF), is a foramen between two spinal Random article vertebrae. Cervical, thoracic, and lumbar Donate to Wikipedia vertebrae all have intervertebral foramina. Wikipedia store The foramina, or openings, are present between Interaction every pair of vertebrae in these areas. A number Help of structures pass through the foramen. These are About Wikipedia the root of each spinal nerve, dorsal root ganglion, Community portal the spinal artery of the segmental artery, Recent changes communicating veins between the internal and Contact page external plexuses, recurrent meningeal (sinu- Tools vertebral) nerves, and transforaminal ligaments. What links here When the spinal vertebrae are articulated with Related changes each other the bodies form a strong pillar for the Upload file support of the head and trunk, and the vertebral Special pages Permanent link foramen constitutes a canal for the protection of Page information the medulla spinalis (spinal cord). Peculiar thoracic vertebrae. Intervertebral foramina are indicated by arrows. Wikidata item The size of the foramina is variable due to Cite this page placement, pathology, spinal loading, and posture. Print/export Foramina can be occluded by arthritic Create a book degenerative changes and space-occupying Download as PDF lesions like tumors, metastases and spinal disc Printable version herniations. Languages Specifically the intervertebral foramen is bound by Deutsch the superior notch of the adjacent vertebra, the Español inferior notch of the vertebra, the vertebral body, and facet joints on the transverse process of the فارسی Français vertebra. -
Bones, Part 1: the Axial Skeleton
PowerPoint® Lecture Slides The Skeleton prepared by Leslie Hendon University of Alabama, Birmingham • Consists of: • Bones, cartilage, joints, and ligaments Composed of 206 named bones grouped into C H A P T E R • 7 two divisions Part 1 • Axial skeleton (80 bones) Bones, Part 1: • Appendicular skeleton (126 bones) The Axial Skeleton Copyright © 2011 Pearson Education, Inc. Copyright © 2011 Pearson Education, Inc. The Axial Skeleton(in green) The Axial Skeleton Cranium Skull Cranium Facial bones Bones of Clavicle pectoral Clavicle girdle • Formed from 80 Thoracic cage Scapula (ribs and Scapula Sternum sternum) Upper named bones Rib limb Humerus Rib Humerus Vertebra • Consists of skull, Vertebral Vertebra Radius column Radius Ulna Ulna vertebral column, Sacrum Carpals Bones of and bony thorax pelvic girdle Carpals Phalanges Phalanges Metacarpals Metacarpals Femur Femur Patella Lower Tibia limb Tibia Fibula Fibula Tarsals Metatarsals (a) Anterior view Phalanges (b) Posterior view Copyright © 2011 Pearson Education, Inc. Figure 7.1a Copyright © 2011 Pearson Education, Inc. Figure 7.1b The Skull The Cranium • Formed by cranial and facial bones • Is the body’s most complex bony structure Frontal bone Parietal bone Glabella • Formed by cranial and facial bones Squamous part Frontonasal suture of frontal bone Supraorbital foramen Nasal bone (notch) • The cranium Sphenoid bone Supraorbital margin (greater wing) Superior orbital fissure • Encloses and protects brain Temporal bone Optic canal Ethmoid bone Inferior orbital fissure Lacrimal bone • Provides attachment for head and neck Zygomatic bone Middle nasal concha muscles Infraorbital foramen Ethmoid Perpendicular plate bone Maxilla Inferior nasal concha Vomer Mandible Mental foramen Mental protuberance (a) Anterior view of skull Copyright © 2011 Pearson Education, Inc.