An Investigation Into the Ecology and Distribu
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
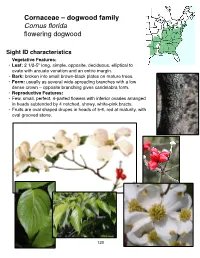
Cornaceae – Dogwood Family Cornus Florida Flowering Dogwood
Cornaceae – dogwood family Cornus florida flowering dogwood Sight ID characteristics Vegetative Features: • Leaf: 2 1/2-5" long, simple, opposite, deciduous, elliptical to ovate with arcuate venation and an entire margin. • Bark: broken into small brown-black plates on mature trees. • Form: usually as several wide-spreading branches with a low dense crown – opposite branching gives candelabra form. • Reproductive Features: • Few, small, perfect, 4-parted flowers with inferior ovaries arranged in heads subtended by 4 notched, showy, white-pink bracts. • Fruits are oval shaped drupes in heads of 5-6, red at maturity, with oval grooved stone. 123 NOTES AND SKETCHES 124 Cornaceae – dogwood family Cornus nuttallii Pacific dogwood Sight ID characteristics Vegetative Features: • Leaf: 2 1/2-4 1/2" long, simple, opposite, deciduous, ovate- elliptical with arcuate venation, margin may be sparsely toothed or entire. • Bark: dark and broken into small plates at maturity. • Form: straight trunk and narrow crown in forested conditions, many-trunked and bushy in open. • Reproductive Features: • Many yellowish-green, small, perfect, 4-parted flowers with inferior ovaries arranged in dense in heads, subtended by 4-7 showy white- pink, petal-like bracts - not notched at the apex. • Fruits are drupes in heads of 30-40, red at maturity and they have smooth stones. 125 NOTES AND SKETCHES 126 Cornaceae – dogwood family Cornus sericea red-osier dogwood Sight ID characteristics Vegetative Features: • Leaf: 2-4" long, simple, opposite, deciduous and somewhat narrow ovate-lanceolate with entire margin. • Twig: bright red, sometimes green splotched with red, white pith. • Bark: red to green with numerous lenticels; later developing larger cracks and splits and turning light brown. -

Chetco Bar BAER Specialist Reports
Chetco Bar BAER Specialist Reports Burned Area Emergency Response Soil Resource Assessment Chetco Bar Fire OR-RSF-000326 Rogue River-Siskiyou National Forest October 2017 Lizeth Ochoa – BAER Team Soil Scientist USFS, Rogue River-Siskiyou NF [email protected] Kit MacDonald – BAER Team Soil Scientist USFS, Coconino and Kaibab National Forests [email protected] 1 EXECUTIVE SUMMARY The Chetco Bar fire occurred on 191,197 acres on the Gold Beach and Wild Rivers Ranger District of the Rogue River-Siskiyou National Forest, BLM lands, and other ownerships in southwestern Oregon. Approximately 170,321 acres of National Forest System (NFS) land, 6,746 acres of BLM land and 14,130 acres of private land were affected by this wildfire. Within the fire perimeter, approximately 14,012 acres burned at high soil burn severity, 64,545 acres burned at moderate soil burn severity, 76,613 acres burned at low soil burn severity, and 36,027 remain unburned. On NFS-managed lands, 10,684 acres burned at high soil burn severity, 58,784 acres burned at moderate soil burn severity, 70,201 acres burned at low soil burn severity and 30,642 acres remain unburned or burned at very low soil burn severity (Figure 1). The Chetco Bar fire burned area is characterized as steep, rugged terrain, with highly dissected slopes and narrow drainages. Dominant surficial geology is metamorphosed sedimentary and volcanic rocks, peridotite and other igneous rocks. Peridotite has been transformed into serpentine through a process known as serpentinization. This transformation is the result of hydration and metamorphic transformation of ultramafic (high iron and magnesium) rocks. -

Likely to Have Habitat Within Iras That ALLOW Road
Item 3a - Sensitive Species National Master List By Region and Species Group Not likely to have habitat within IRAs Not likely to have Federal Likely to have habitat that DO NOT ALLOW habitat within IRAs Candidate within IRAs that DO Likely to have habitat road (re)construction that ALLOW road Forest Service Species Under NOT ALLOW road within IRAs that ALLOW but could be (re)construction but Species Scientific Name Common Name Species Group Region ESA (re)construction? road (re)construction? affected? could be affected? Bufo boreas boreas Boreal Western Toad Amphibian 1 No Yes Yes No No Plethodon vandykei idahoensis Coeur D'Alene Salamander Amphibian 1 No Yes Yes No No Rana pipiens Northern Leopard Frog Amphibian 1 No Yes Yes No No Accipiter gentilis Northern Goshawk Bird 1 No Yes Yes No No Ammodramus bairdii Baird's Sparrow Bird 1 No No Yes No No Anthus spragueii Sprague's Pipit Bird 1 No No Yes No No Centrocercus urophasianus Sage Grouse Bird 1 No Yes Yes No No Cygnus buccinator Trumpeter Swan Bird 1 No Yes Yes No No Falco peregrinus anatum American Peregrine Falcon Bird 1 No Yes Yes No No Gavia immer Common Loon Bird 1 No Yes Yes No No Histrionicus histrionicus Harlequin Duck Bird 1 No Yes Yes No No Lanius ludovicianus Loggerhead Shrike Bird 1 No Yes Yes No No Oreortyx pictus Mountain Quail Bird 1 No Yes Yes No No Otus flammeolus Flammulated Owl Bird 1 No Yes Yes No No Picoides albolarvatus White-Headed Woodpecker Bird 1 No Yes Yes No No Picoides arcticus Black-Backed Woodpecker Bird 1 No Yes Yes No No Speotyto cunicularia Burrowing -

Mountain Laurel Or Rhododendron? Recreation Area Vol
U.S. Dept. of the Interior National Park Service . Spanning the Gap The newsletter of Delaware Water Gap National Mountain Laurel or Rhododendron? Recreation Area Vol. 7 No. 2 Summer 1984 These two members of the plant kingdom are often confused. Especially before they blossom, they resemble each other with their bushy appearance and long, dark-green leaves. Both plants ARE shrubs, and they are often seen growing together. Another thing they have in common: although some species have not been proven dangerous, it is well to consider ALL members of the laurel- (Above) Rhodendron. rhododendron-azalea group of plants as potentially toxic. The rhododendron (rhododenron maximum) is undoubtedly the most conspicuous understory plant in the mountain forests of Pennsylvania. In ravines and hollows and along shaded watercourses it often grows so luxuriantly as to form almost impenetrable tangles. In June or early July, large clusters of showy blossoms, ranging from white to light rosy- pink, appear at the ends of the branchlets. People travel many miles to witness the magnificent show which is provided by the rhododendron at flowering time. In the southern Appalachians the plant grows much larger than it ordinarily does in this area, often becoming a tree 25 feet or more in height. The dense thickets of rhododendron in the stream valleys and on the lower slopes of our mountains are favorite yarding grounds for deer when the Rhodendron. snows become deep in the wintertime. The dark- green, leathery leaves react to subfreezing temperatures by bending downward and rolling into a tight coil. Although the shrub is often browsed to excess, it has little or no nutritional value. -

Coptis Trifolia Conservation Assessment
CONSERVATION ASSESSMENT for Coptis trifolia (L.) Salisb. Originally issued as Management Recommendations December 1998 Marty Stein Reconfigured-January 2005 Tracy L. Fuentes USDA Forest Service Region 6 and USDI Bureau of Land Management, Oregon and Washington CONSERVATION ASSESSMENT FOR COPTIS TRIFOLIA Table of Contents Page List of Tables ................................................................................................................................. 2 List of Figures ................................................................................................................................ 2 Summary........................................................................................................................................ 4 I. NATURAL HISTORY............................................................................................................. 6 A. Taxonomy and Nomenclature.......................................................................................... 6 B. Species Description ........................................................................................................... 6 1. Morphology ................................................................................................................... 6 2. Reproductive Biology.................................................................................................... 7 3. Ecological Roles ............................................................................................................. 7 C. Range and Sites -

Arctostaphylos Hispidula, Gasquet Manzanita
Conservation Assessment for Gasquet Manzanita (Arctostaphylos hispidula) Within the State of Oregon Photo by Clint Emerson March 2010 U.S.D.A. Forest Service Region 6 and U.S.D.I. Bureau of Land Management Interagency Special Status and Sensitive Species Program Author CLINT EMERSON is a botanist, USDA Forest Service, Rogue River-Siskiyou National Forest, Gold Beach and Powers Ranger District, Gold Beach, OR 97465 TABLE OF CONTENTS Disclaimer 3 Executive Summary 3 List of Tables and Figures 5 I. Introduction 6 A. Goal 6 B. Scope 6 C. Management Status 7 II. Classification and Description 8 A. Nomenclature and Taxonomy 8 B. Species Description 9 C. Regional Differences 9 D. Similar Species 10 III. Biology and Ecology 14 A. Life History and Reproductive Biology 14 B. Range, Distribution, and Abundance 16 C. Population Trends and Demography 19 D. Habitat 21 E. Ecological Considerations 25 IV. Conservation 26 A. Conservation Threats 26 B. Conservation Status 28 C. Known Management Approaches 32 D. Management Considerations 33 V. Research, Inventory, and Monitoring Opportunities 35 Definitions of Terms Used (Glossary) 39 Acknowledgements 41 References 42 Appendix A. Table of Known Sites in Oregon 45 2 Disclaimer This Conservation Assessment was prepared to compile existing published and unpublished information for the rare vascular plant Gasquet manzanita (Arctostaphylos hispidula) as well as include observational field data gathered during the 2008 field season. This Assessment does not represent a management decision by the U.S. Forest Service (Region 6) or Oregon/Washington BLM. Although the best scientific information available was used and subject experts were consulted in preparation of this document, it is expected that new information will arise. -

Apn80: Northern Spruce
ACID PEATLAND SYSTEM APn80 Northern Floristic Region Northern Spruce Bog Black spruce–dominated peatlands on deep peat. Canopy is often sparse, with stunted trees. Understory is dominated by ericaceous shrubs and fine- leaved graminoids on high Sphagnum hummocks. Vegetation Structure & Composition Description is based on summary of vascular plant data from 84 plots (relevés) and bryophyte data from 17 plots. l Moss layer consists of a carpet of Sphag- num with moderately high hummocks, usually surrounding tree bases, and weakly developed hollows. S. magellanicum dominates hum- mocks, with S. angustifolium present in hollows. High hummocks of S. fuscum may be present, although they are less frequent than in open bogs and poor fens. Pleurozium schre- beri is often very abundant and forms large mats covering drier mounds in shaded sites. Dicranum species are commonly interspersed within the Pleurozium mats. l Forb cover is minimal, and may include three-leaved false Solomon’s seal (Smilacina trifolia) and round-leaved sundew (Drosera rotundifolia). l Graminoid cover is 5-25%. Fine-leaved graminoids are most important and include three-fruited bog sedge (Carex trisperma) and tussock cottongrass (Eriophorum vagi- natum). l Low-shrub layer is prominent and dominated by ericaceous shrubs, particularly Lab- rador tea (Ledum groenlandicum), which often has >25% cover. Other ericads include bog laurel (Kalmia polifolia), small cranberry (Vaccinium oxycoccos), and leatherleaf (Chamaedaphne calyculata). Understory trees are limited to scattered black spruce. l Canopy is dominated by black spruce. Trees are usually stunted (<30ft [10m] tall) with 25-75% cover . Some sites have scattered tamaracks in addition to black spruce. -

Kalmia Procumbens (L.) Gift & Kron Synonym: Loiseleuria Procumbens (L.) Desv
Kalmia procumbens (L.) Gift & Kron synonym: Loiseleuria procumbens (L.) Desv. alpine azalea Ericaceae - heath family status: State Threatened, BLM sensitive, USFS sensitive rank: G5 / S1 General Description: Adapted from Douglas et al. (1998-2002): A diffusely branched, dwarf evergreen shrub, trailing on the ground without rooting at the nodes. Stems 5-30 cm long, hairless or minutely hairy. Leaves with partially clasping stalks, opposite; blades leathery, bright green, narrowly elliptic, 3-8 x 1-3 mm, hairless above, short hairs below, with revolute margins. Floral Characteristics: Flowers 2-6, light to deep pink, clustered in axils of upper stem leaves, erect on short stalks 1-10 mm long, straight or curved. Corolla bell-shaped, 5-lobed, 4-5 mm long. Calyx red, 1.5-2.5 mm long, hairless, persistent. Stamens 5, anthers unawned, opening full length by slits. Fruits: Ovoid capsules 3-5 x 2-3 mm. Identifiable July to August. Identification Tips: Phyllodoce species are closely related to Kalmia, but they have alternate, linear leaves 6-16 mm long, usually 10 stamens, and anthers that open by two terminal pores. Cassiope species have smaller leaves (less than 5 mm long) lacking revolute margins, awned anthers that open by terminal pores, and usually nodding flowers. K. procumbens differs from K. microphylla (two varieties occur in Washington) in having capsules with 2-3 chambers, smaller petals ( 3-5 mm vs. 7-9 mm) cleft half their length, and 5 stamens. K. © Margaret Williams microphylla has capsules with 5 chambers, petals that are only shallowly cleft, and 10 stamens. Range: Circumpolar. -

U.S. GEOLOGICAL SURVEY PROFESSIONAL PAPER 1378 Deformation of the Josephine Peridotite, California and Oregon
U.S. GEOLOGICAL SURVEY PROFESSIONAL PAPER 1378 Deformation of the Josephine Peridotite, California and Oregon By JAMES G. EVANS U.S. GEOLOGICAL SURVEY PROFESSIONAL PAPER 137 UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1987 DEPARTMENT OF THE INTERIOR DONALD PAUL HODEL, Secretary U.S. GEOLOGICAL SURVEY Dallas L. Peck, Director Library of Congress Cataloging in Publication Data Evans, James George, 1938- Deformation of the Josephine Peridotite, California and Oregon. (U.S. Geological Survey Professional Paper 1378) Bibliography: p. 44-45 Supt. of Docs. No.: I 19.16:1378 1. Rock deformation Klamath Mountains (Calif, and Oreg.). 2. Geology, stratigraphic Mesozoic. 3. Peridotite Klamath Mountains (Calif, and Oreg.). 4. Geology Klamath Mountains (Calif, and Oreg.). I. Title. II. Series: United States. Geological Survey. Professional Paper 1378. QE604.E88 1986 551.8'09794'21 87-600231 For sale by the Books and Open-File Reports Section, U.S. Geological Survey, Federal Center, Box 25425, Denver, CO 80225 CONTENTS Page Abstract __________________________________ 1 Structural analysis Continued Introduction ______________________________ 2 Mesoscopic fabric Continued Geologic setting__________________ __________ 2 Fabric of the Vulcan Peak area __ ____________ 32 Josephine Peridotite_________________________ 5 Petrofabric studies ___________________________ 33 Structural analysis __________________________ 7 Introduction ____________________________ 33 Introduction ___________________________ 7 Description of microfabrics ____________________ -

Rare Plant Survey of San Juan Public Lands, Colorado
Rare Plant Survey of San Juan Public Lands, Colorado 2005 Prepared by Colorado Natural Heritage Program 254 General Services Building Colorado State University Fort Collins CO 80523 Rare Plant Survey of San Juan Public Lands, Colorado 2005 Prepared by Peggy Lyon and Julia Hanson Colorado Natural Heritage Program 254 General Services Building Colorado State University Fort Collins CO 80523 December 2005 Cover: Imperiled (G1 and G2) plants of the San Juan Public Lands, top left to bottom right: Lesquerella pruinosa, Draba graminea, Cryptantha gypsophila, Machaeranthera coloradoensis, Astragalus naturitensis, Physaria pulvinata, Ipomopsis polyantha, Townsendia glabella, Townsendia rothrockii. Executive Summary This survey was a continuation of several years of rare plant survey on San Juan Public Lands. Funding for the project was provided by San Juan National Forest and the San Juan Resource Area of the Bureau of Land Management. Previous rare plant surveys on San Juan Public Lands by CNHP were conducted in conjunction with county wide surveys of La Plata, Archuleta, San Juan and San Miguel counties, with partial funding from Great Outdoors Colorado (GOCO); and in 2004, public lands only in Dolores and Montezuma counties, funded entirely by the San Juan Public Lands. Funding for 2005 was again provided by San Juan Public Lands. The primary emphases for field work in 2005 were: 1. revisit and update information on rare plant occurrences of agency sensitive species in the Colorado Natural Heritage Program (CNHP) database that were last observed prior to 2000, in order to have the most current information available for informing the revision of the Resource Management Plan for the San Juan Public Lands (BLM and San Juan National Forest); 2. -

Agroforestry News Index Vol 1 to Vol 22 No 2
Agroforestry News Index Vol 1 to Vol 22 No 2 2 A.R.T. nursery ..... Vol 2, No 4, page 2 Acorns, edible from oaks ..... Vol 5, No 4, page 3 Aaron, J R & Richards: British woodland produce (book review) ..... Acorns, harvesting ..... Vol 5, No 4, Vol 1, No 4, page 34 page 3 Abies balsamea ..... Vol 8, No 2, page Acorns, nutritional composition ..... 31 Vol 5, No 4, page 4 Abies sibirica ..... Vol 8, No 2, page 31 Acorns, removing tannins from ..... Vol 5, No 4, page 4 Abies species ..... Vol 19, No 1, page 13 Acorns, shelling ..... Vol 5, No 4, page 3 Acca sellowiana ..... Vol 9, No 3, page 4 Acorns, utilisation ..... Vol 5, No 4, page 4 Acer macrophyllum ..... Vol 16, No 2, page 6 Acorus calamus ..... Vol 8, No 4, page 6 Acer pseudoplatanus ..... Vol 3, No 1, page 3 Actinidia arguta ..... Vol 1, No 4, page 10 Acer saccharum ..... Vol 16, No 1, page 3 Actinidia arguta, cultivars ..... Vol 1, No 4, page 14 Acer saccharum - strawberry agroforestry system ..... Vol 8, No 1, Actinidia arguta, description ..... Vol page 2 1, No 4, page 10 Acer species, with edible saps ..... Vol Actinidia arguta, drawings ..... Vol 1, 2, No 3, page 26 No 4, page 15 Achillea millefolium ..... Vol 8, No 4, Actinidia arguta, feeding & irrigaton page 5 ..... Vol 1, No 4, page 11 3 Actinidia arguta, fruiting ..... Vol 1, Actinidia spp ..... Vol 5, No 1, page 18 No 4, page 13 Actinorhizal plants ..... Vol 3, No 3, Actinidia arguta, nurseries page 30 supplying ..... Vol 1, No 4, page 16 Acworth, J M: The potential for farm Actinidia arguta, pests and diseases forestry, agroforestry and novel tree .... -

Proceedings of the United States National Museum
A REVISION OF CERTAIN SPECIES OF PLANTS OF THE GENUS ANTENNARIA. By Elias Nelson. Scieniific Ai<L U. S. Department of A(jrietiJliire. Recent study of the genus Anton niiria in North America has been (djVHa, A. contined almost entirely to the o-roup represented by .1. These, as dioka, and A. plantaglnifolia of Gray's Synoptical Flora. treated by Dr. Gray, have proved to be ac^gregates. The names .1. plants, and (djnna and .i. dloJca were originally applied to Old World referal)le to those species. it appears that no American specimens are had Prior to 1897 only live species of the North American continent Rich- been described in this group, viz, ^L planfaghiifolia (Linna?us) pJanta- ardson, A. monocephala De CandoUe, A. soUtaria Rydberg (.1. Nuttall, and ginifolia monocephala Torrey and Gray), A. parolfdia about A. lahradorica Nuttall. During the last four years, however, 50 additional species have been published. The Antennarias of many parts of North America are as yet little known, and much research Fernald has in connection with the genus is still necessary. Mr, succeeded in arriving at a very satisfactory arrangement of the New England Antennarias. Those of other regions are in need of similar study. of the In this paper an attempt is made at a natural arrangement western and northern species of this group. A tolerably satisfactory synopsis can hardly be expected until more material from this vast imperfectly known; region is at hand. Many of the species are very Especially is a considerable number from their type localities only. variability this true of the far northern species.