Dimorphism in Two New Genera of Devonian Tabulate Corals
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

University of Michigan University Library
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN VOL.XIX, No. 3, pp. 23-36 (7 pls.) JULY 17, 1964 CORALS OF THE TRAVERSE GROUP OF MICHIGAN PART XII, THE SMALL-CELLED SPECIES OF FAVOSITES AND EMMONSIA BY ERWIN C. STUMM and JOHN H. TYLER MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR, MICHIGAN CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Director: LEWIS B. KELLUM The series of contributions from the Museum of Paleontology is a medium for the publication of papers based chiefly upon the collections in the Museum. When the number of pages issued is sufficient to make a volume, a title page and a table of contents will be sent to libraries on the mailing list, and to individuals upon request. A list of the separate papers may also be obtained. Correspondence should be directed to the Museum of Paleontology, The University of Michigan, Ann Arbor, Michigan. VOLS.11-XVIII. Parts of volumes may be obtained if available. VOLUMEXIX 1. Silicified Trilobites from the Devonian Jeffersonville Limestone at the Falls of the Ohio, by Erwin C. Stumm. Pages 1-14, with 3 plates. 2. Two Gastropods from the Lower Cretaceous (Albian) of Coahuila, Mexico, by Lewis B. Kellum and Kenneth E. Appelt. Pages 14-22. 3. Corals of the Traverse Group of Michigan, Part XII, The Small-celled Species of Favosites and Emmonsia, by Erwin C. Stumm and John H. Tyler. Pages 23-36, with 7 plates. VOL. XIX, NO.3, pp. 23-36 (7 pls.) JULY 17, 1964 CORALS OF THE TRAVERSE GROUP OF MICHIGAN PART XII, THE SMALL-CELLED SPECIES OF FAVOSZTES AND EMMONSIA1 BY ERWIN C. -

United States
DEPARTMENT OF THE INTERIOR BULLETIN OF THE UNITED STATES ISTo. 146 WASHINGTON GOVERNMENT Pit IN TING OFFICE 189C UNITED STATES GEOLOGICAL SURVEY CHAKLES D. WALCOTT, DI11ECTOK BIBLIOGRAPHY AND INDEX NORTH AMEEICAN GEOLOGY, PALEONTOLOGY, PETEOLOGT, AND MINERALOGY THE YEA.R 1895 FEED BOUGHTON WEEKS WASHINGTON Cr O V E U N M K N T P K 1 N T I N G OFFICE 1890 CONTENTS. Page. Letter of trail smittal...... ....................... .......................... 7 Introduction.............'................................................... 9 List of publications examined............................................... 11 Classified key to tlio index .......................................... ........ 15 Bibliography ............................................................... 21 Index....................................................................... 89 LETTER OF TRANSMITTAL DEPARTMENT OF THE INTEEIOE, UNITED STATES GEOLOGICAL SURVEY, DIVISION OF GEOLOGY, Washington, D. 0., June 23, 1896. SIR: I have the honor to transmit herewith the manuscript of a Bibliography and Index of North American Geology, Paleontology, Petrology, and Mineralogy for the year 1895, and to request that it be published as a bulletin of the Survey. Very respectfully, F. B. WEEKS. Hon. CHARLES D. WALCOTT, Director United States Geological Survey. 1 BIBLIOGRAPHY AND INDEX OF NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINER ALOGY FOR THE YEAR 1895. By FRED BOUGHTON WEEKS. INTRODUCTION. The present work comprises a record of publications on North Ameri can geology, paleontology, petrology, and mineralogy for the year 1895. It is planned on the same lines as the previous bulletins (Nos. 130 and 135), excepting that abstracts appearing in regular periodicals have been omitted in this volume. Bibliography. The bibliography consists of full titles of separate papers, classified by authors, an abbreviated reference to the publica tion in which the paper is printed, and a brief summary of the con tents, each paper being numbered for index reference. -

Geological Survey
DEPARTMENT OF THE INTERIOR UNITED STATES GEOLOGICAL SURVEY ISTo. 162 WASHINGTON GOVERNMENT PRINTING OFFICE 1899 UNITED STATES GEOLOGICAL SUEVEY CHAKLES D. WALCOTT, DIRECTOR Olf NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINERALOGY FOR THE YEAR 1898 BY FRED BOUG-HTOISr WEEKS WASHINGTON GOVERNMENT PRINTING OFFICE 1899 CONTENTS, Page. Letter of transmittal.......................................... ........... 7 Introduction................................................................ 9 List of publications examined............................................... 11 Bibliography............................................................... 15 Classified key to the index .................................................. 107 Indiex....................................................................... 113 LETTER OF TRANSMITTAL. DEPARTMENT OF THE INTERIOR, UNITED STATES GEOLOGICAL SURVEY. Washington, D. C., June 30,1899. SIR: I have the honor to transmit herewith the manuscript of a Bibliography and Index o'f North American Geology, Paleontology, Petrology, and Mineralogy for the Year 1898, and to request that it be published as a bulletin of the Survey. Very respectfully, F. B. WEEKS. Hon. CHARLES D. WALCOTT, Director United States Geological Survey. 1 I .... v : BIBLIOGRAPHY AND INDEX OF NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINERALOGY FOR THE YEAR 1898. ' By FEED BOUGHTON WEEKS. INTRODUCTION. The method of preparing and arranging the material of the Bibli ography and Index for 1898 is similar to that adopted for the previous publications 011 this subject (Bulletins Nos. 130,135,146,149, and 156). Several papers that should have been entered in the previous bulletins are here recorded, and the date of publication is given with each entry. Bibliography. The bibliography consists of full titles of separate papers, classified by authors, an abbreviated reference to the publica tion in which the paper is printed, and a brief summary of the con tents, each paper being numbered for index reference. -

CNIDARIA Corals, Medusae, Hydroids, Myxozoans
FOUR Phylum CNIDARIA corals, medusae, hydroids, myxozoans STEPHEN D. CAIRNS, LISA-ANN GERSHWIN, FRED J. BROOK, PHILIP PUGH, ELLIOT W. Dawson, OscaR OcaÑA V., WILLEM VERvooRT, GARY WILLIAMS, JEANETTE E. Watson, DENNIS M. OPREsko, PETER SCHUCHERT, P. MICHAEL HINE, DENNIS P. GORDON, HAMISH J. CAMPBELL, ANTHONY J. WRIGHT, JUAN A. SÁNCHEZ, DAPHNE G. FAUTIN his ancient phylum of mostly marine organisms is best known for its contribution to geomorphological features, forming thousands of square Tkilometres of coral reefs in warm tropical waters. Their fossil remains contribute to some limestones. Cnidarians are also significant components of the plankton, where large medusae – popularly called jellyfish – and colonial forms like Portuguese man-of-war and stringy siphonophores prey on other organisms including small fish. Some of these species are justly feared by humans for their stings, which in some cases can be fatal. Certainly, most New Zealanders will have encountered cnidarians when rambling along beaches and fossicking in rock pools where sea anemones and diminutive bushy hydroids abound. In New Zealand’s fiords and in deeper water on seamounts, black corals and branching gorgonians can form veritable trees five metres high or more. In contrast, inland inhabitants of continental landmasses who have never, or rarely, seen an ocean or visited a seashore can hardly be impressed with the Cnidaria as a phylum – freshwater cnidarians are relatively few, restricted to tiny hydras, the branching hydroid Cordylophora, and rare medusae. Worldwide, there are about 10,000 described species, with perhaps half as many again undescribed. All cnidarians have nettle cells known as nematocysts (or cnidae – from the Greek, knide, a nettle), extraordinarily complex structures that are effectively invaginated coiled tubes within a cell. -

Geology of the Hurley West Quadrangle Grant County New Mexico
Geology of the Hurley West Quadrangle Grant County New Mexico By WALDEN P. PRATT CONTRIBUTIONS TO GENERAL GEOLOGY GEOLOGICAL SURVEY BULLETIN 1241-E A study of part of the Silver City mining region, with emphasis on Paleozoic stratigraphy and on early Tertiary intrusion and faulting UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1967 UNITED STATES DEPARTMENT OF THE INTERIOR STEWART L. UDALL, Secretary GEOLOGICAL SURVEY William T. Pecora, Director For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 CONTENTS Page. Abstract ...___.___._._-_______._.___________._______._.._______ El Introduction. _____________________________________________________ 3 Location and geologic significance of area.-_-__---_______________ 3 Geography. ___ _ __-- _ 5 Purpose of investigation._______________________________________ 5 Previous work__-----------------_-__-_-____________________ 6 Fieldwork and acknowledgments________________________________ 8 Geologic formations-____-_-----__--_-__--__--____-_-_______________ 8 Precambrian rockl_____-----_---_-_____________________________ 9 Paleozoic rocks_______-------------_----_______________________ 10 Cambrian and Ordovician Systems, Bliss Sandstone__________ 10 Ordovician System________________________________________ 15 El Paso Dolomite____________________________________ 15 Montoya Group_______________________________________ 18 Distribution. _____________________________________ 19 Subdivision.______________________________________ 19 Second Value Dolomite___________________________ -

Tayside, Central and Fife Tayside, Central and Fife
Detail of the Lower Devonian jawless, armoured fish Cephalaspis from Balruddery Den. © Perth Museum & Art Gallery, Perth & Kinross Council Review of Fossil Collections in Scotland Tayside, Central and Fife Tayside, Central and Fife Stirling Smith Art Gallery and Museum Perth Museum and Art Gallery (Culture Perth and Kinross) The McManus: Dundee’s Art Gallery and Museum (Leisure and Culture Dundee) Broughty Castle (Leisure and Culture Dundee) D’Arcy Thompson Zoology Museum and University Herbarium (University of Dundee Museum Collections) Montrose Museum (Angus Alive) Museums of the University of St Andrews Fife Collections Centre (Fife Cultural Trust) St Andrews Museum (Fife Cultural Trust) Kirkcaldy Galleries (Fife Cultural Trust) Falkirk Collections Centre (Falkirk Community Trust) 1 Stirling Smith Art Gallery and Museum Collection type: Independent Accreditation: 2016 Dumbarton Road, Stirling, FK8 2KR Contact: [email protected] Location of collections The Smith Art Gallery and Museum, formerly known as the Smith Institute, was established at the bequest of artist Thomas Stuart Smith (1815-1869) on land supplied by the Burgh of Stirling. The Institute opened in 1874. Fossils are housed onsite in one of several storerooms. Size of collections 700 fossils. Onsite records The CMS has recently been updated to Adlib (Axiel Collection); all fossils have a basic entry with additional details on MDA cards. Collection highlights 1. Fossils linked to Robert Kidston (1852-1924). 2. Silurian graptolite fossils linked to Professor Henry Alleyne Nicholson (1844-1899). 3. Dura Den fossils linked to Reverend John Anderson (1796-1864). Published information Traquair, R.H. (1900). XXXII.—Report on Fossil Fishes collected by the Geological Survey of Scotland in the Silurian Rocks of the South of Scotland. -
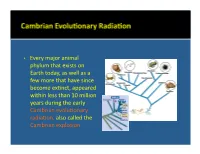
• Every Major Animal Phylum That Exists on Earth Today, As Well As A
• Every major animal phylum that exists on Earth today, as well as a few more that have since become ex:nct, appeared within less than 10 million years during the early Cambrian evolu:onary radiaon, also called the Cambrian explosion. • Phylum Pro:sta includes a diverse group of eukaryo:c microorganisms which are mostly unicellular. They are not necessarily primi:ve, although some are. Only some have a skeleton that can be preserved as a fossil. • Foraminifera and radiolaria are the most important examples. Coccolithophores, diatoms, and dinoflagellates are also examples. • Foraminifera are marine organisms that may be either planktonic, living in the water column where they float at various levels, or benthic, living on or within the seafloor sediment. • They form relavely large (0.1 mm to 5 cm) porous internal skeletons called tests through which project strands of living cytoplasm. The single- to mul:-chambered tests may be composed of organic maer, agglu:nated sand grains or sponge spicules, or calcium carbonate. • Benthic foraminifera • Planktonic foraminifera (Cambrian to Recent). (Jurassic to Recent). Oligocene Eocene • Radiolaria are planktonic marine organisms that tend to live in relavely cold water. • Their tests, oNen delicate and elaborate, are made of silica and presently accumulate on the floors of the parts of oceans that are deeper than those where foraminiferal tests are accumulang. • Radiolaria (Cambrian to • Radiolaria (Cambrian to Recent). Recent). Paleocene Radiolaria • Coccolithophores (Triassic to Recent) are extremely small marine planktonic photosynthe:c unicellular algae that are enclosed by plates of low-Mg calcite called coccoliths. Cretaceous White Cliffs of Dover, England • Diatoms (Jurassic to Recent) are small freshwater and marine planktonic photosynthe:c unicellular algae that are enclosed by a cell wall made of silica called a frustule. -

Back Matter (PDF)
Index Page numbers in italic denote Figures. Page numbers in bold denote Tables. Acadian Orogeny 224 Ancyrodelloides delta biozone 15 Acanthopyge Limestone 126, 128 Ancyrodelloides transitans biozone 15, 17,19 Acastella 52, 68, 69, 70 Ancyrodelloides trigonicus biozone 15, 17,19 Acastoides 52, 54 Ancyrospora 31, 32,37 Acinosporites lindlarensis 27, 30, 32, 35, 147 Anetoceras 82 Acrimeroceras 302, 313 ?Aneurospora 33 acritarchs Aneurospora minuta 148 Appalachian Basin 143, 145, 146, 147, 148–149 Angochitina 32, 36, 141, 142, 146, 147 extinction 395 annulata Events 1, 2, 291–344 Falkand Islands 29, 30, 31, 32, 33, 34, 36, 37 comparison of conodonts 327–331 late Devonian–Mississippian 443 effects on fauna 292–293 Prague Basin 137 global recognition 294–299, 343 see also Umbellasphaeridium saharicum limestone beds 3, 246, 291–292, 301, 308, 309, Acrospirifer 46, 51, 52, 73, 82 311, 321 Acrospirifer eckfeldensis 58, 59, 81, 82 conodonts 329, 331 Acrospirifer primaevus 58, 63, 72, 74–77, 81, 82 Tafilalt fauna 59, 63, 72, 74, 76, 103 ammonoid succession 302–305, 310–311 Actinodesma 52 comparison of facies 319, 321, 323, 325, 327 Actinosporites 135 conodont zonation 299–302, 310–311, 320 Acuticryphops 253, 254, 255, 256, 257, 264 Anoplia theorassensis 86 Acutimitoceras 369, 392 anoxia 2, 3–4, 171, 191–192, 191 Acutimitoceras (Stockumites) 357, 359, 366, 367, 368, Hangenberg Crisis 391, 392, 394, 401–402, 369, 372, 413 414–417, 456 agnathans 65, 71, 72, 273–286 and carbon cycle 410–413 Ahbach Formation 172 Kellwasser Events 237–239, 243, 245, 252 -

Synoptic Taxonomy of Major Fossil Groups
APPENDIX Synoptic Taxonomy of Major Fossil Groups Important fossil taxa are listed down to the lowest practical taxonomic level; in most cases, this will be the ordinal or subordinallevel. Abbreviated stratigraphic units in parentheses (e.g., UCamb-Ree) indicate maximum range known for the group; units followed by question marks are isolated occurrences followed generally by an interval with no known representatives. Taxa with ranges to "Ree" are extant. Data are extracted principally from Harland et al. (1967), Moore et al. (1956 et seq.), Sepkoski (1982), Romer (1966), Colbert (1980), Moy-Thomas and Miles (1971), Taylor (1981), and Brasier (1980). KINGDOM MONERA Class Ciliata (cont.) Order Spirotrichia (Tintinnida) (UOrd-Rec) DIVISION CYANOPHYTA ?Class [mertae sedis Order Chitinozoa (Proterozoic?, LOrd-UDev) Class Cyanophyceae Class Actinopoda Order Chroococcales (Archean-Rec) Subclass Radiolaria Order Nostocales (Archean-Ree) Order Polycystina Order Spongiostromales (Archean-Ree) Suborder Spumellaria (MCamb-Rec) Order Stigonematales (LDev-Rec) Suborder Nasselaria (Dev-Ree) Three minor orders KINGDOM ANIMALIA KINGDOM PROTISTA PHYLUM PORIFERA PHYLUM PROTOZOA Class Hexactinellida Order Amphidiscophora (Miss-Ree) Class Rhizopodea Order Hexactinosida (MTrias-Rec) Order Foraminiferida* Order Lyssacinosida (LCamb-Rec) Suborder Allogromiina (UCamb-Ree) Order Lychniscosida (UTrias-Rec) Suborder Textulariina (LCamb-Ree) Class Demospongia Suborder Fusulinina (Ord-Perm) Order Monaxonida (MCamb-Ree) Suborder Miliolina (Sil-Ree) Order Lithistida -

4. Arbeitstreffen Deutschsprachiger Echinodermenforscher Wien, Oktober 08
4. Arbeitstreffen deutschsprachiger Echinodermenforscher Wien, Oktober 08 4. Arbeitstreffen deutschsprachiger Echinodermenforscher, Wien, Okt. 08 4th Workshop of German & Austrian Echinoderm Research 4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research 24.-26. October 2008 ABSTRACTS Andreas Kroh & Brigitta Schmid Naturhistorisches Museum Wien 4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research Impressum Eigentümer, Herausgeber und Verleger: © 2008 Natuhistorisches Museum Wien Alle Rechte vorbehalten. Für den Inhalt der Beiträge sind die Autoren verantwortlich. Redaktion: Naturhistorisches Museum Wien Burgring 7, A-1010 Wien, Österreich (Austria) Tel.: +43 (1) 521 77 / 576 (Kroh); +43 (1) 521 77 / 564 (Schmid) Fax: +43 (1) 521 77 / 459 e-mail: [email protected] e-mail: [email protected] Redaktionelle Bearbeitung (EDV, Layout, Grafik): Andreas Kroh Umschlagentwurf: Andreas Kroh Titelbild: Coelopleurus (Keraiophorus) exquisitus COPPARD & SCHULTZ, 2006; Neukaledonien (NHMW 2005z0277/0016; Photo von Alice SCHUMACHER, NHM Wien). 24.-26. October 2008 iii 4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research Inhalt Encrinus sp. cf. E. robustus (Crinoidea, Encrinidae) aus dem Unteren Muschelkalk von Niedersachsen (Poster) Ulrich Bielert, Friedrich Bielert & Hans Hagdorn...........................................1 Classification of pre- and postmortem -

PREFACE a Graduate Thesis Often Represents a Student's First Attempt at Reporting Extensively on a Specific Research Problem
PREFACE A graduate thesis often represents a student's first attempt at reporting extensively on a specific research problem. The results are seldom published and those unpublished, too often are little known or used. Nevertheless, many if not most of them are sources of potentially significant information. The purpose of this circular is to compile a handy reference to this field of geologic information. Most thesis titles pertain to Michigan and surrounding Great Lakes. However, a few titles were included from adjacent states where this information may be useful in interpreting Michigan geology. The list is organized by author, each thesis assigned a number, cross-referenced to the subject index inside rear cover. The information was gathered primarily by canvassing collegiate geology departments, without whose cooperation this circular would not have been possible. The earliest thesis reported was written in 1895. Revisions are contemplated so any comments or additional data suggested will be appreciated. Prof. B. John Chronic, University of Colorado, has compiled "Bibliography of Theses in Geology" containing over 11,000 titles. University Microfilms, Ann Arbor, Michigan has been microfilming Ph.D. theses since 1938, listed monthly in "Dissertation Abstracts". Ph.D. Geological Survey theses titles appear in the monthly "Abstracts of North CIRCULAR 7 American Geology", U.S. Geological Survey. Lansing Edward A. Kirkby, Geologist INDEX TO MICHIGAN GEOLOGIC THESES May, 1967 Geological Survey Division by E. A. Kirkby Petrography and Petrology (including insoluble residues, petrofabrics, and sediments): Lansing 1967 14, 15, 25, 27, 30, 50, 56, 57, 58, 68, 69, 73, 74, 76, 79, 80, 82, 84, 89, 91, 93, 94, 106, 107, 108, 113, STATE OF MICHIGAN 118, 123, 126, 132, 133, 134, 135, 137, 140, 143, George Romney, Governor 146, 151, 156, 171, 176, 184, 185, 188, 198, 200, DEPARTMENT OF CONSERVATION 216, 218, 224, 226, 227, 237, 243, 246, 253, 259, Ralph A. -

Corals of Falls of the Ohio State Park
The Rugose Corals Tabulate Corals A Devonian coral garden Tabulae Rugose corals get their name because the exterior of Tabulate corals are always colonial. There are 212 species of corals many of their forms has a wrinkly appearance. They are Their name is derived from a flat shelf known from the fossil beds at the often called “horn corals” because their form may resemble (tabula) that separates individuals Falls of the Ohio. Not all occur in CORALS the horn of a cow or goat. In fact, the largest horn coral vertically in the colony (figure 6). The the coral beds; some may be found (Siphonophrentis elongata, figure 1) was referred to as a individual corallites that make up the in the slightly higher (younger) at “petrified buffalo horn” by colony are typically 0.5 - 8 mm wide. rock layers. Additional studies may Figure 11. Underside Falls of the Ohio settlers due to its shape. Figure 6. Despite the small individuals, these reveal more or fewer species. It takes of Alveolites coral Horn corals show a wide corals can be quite large. The most experience to identify corals with State Park variation in form, although common coral is a form called Favosites (Emmonsia). accuracy. The random orientation of many of the solitary external form is not a Emmonsia was given its own genus name, but some rugose corals make identification even more difficult. Alan Goldstein, Interpretive Naturalist distinguishing feature for specialists consider it to be a sub-genus of Favosites. These Look for evidence of Devonian hurricanes. Can you find identification.