Odonata Occurrence in Caves: Active Or Accidentals? a New Case Study
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1 June 2021 Researchgate: Researchgate.Net/Profile
DAVID OUTOMURO PRIEDE, PH.D. CURRICULUM VITAE June 2021 Researchgate: researchgate.net/profile/David_Outomuro ORCID: orcid.org/0000-0002-1296-7273 EDUCATION Ph.D. 2011 University of Oviedo, Spain (Biology). Summa cum laude. (Dr. Francisco J. Ocharan) B.S. 2005 University of Oviedo, Spain (Biology). Valedictorian. PROFESSIONAL EXPERIENCE Aug 2017- Aug 2021 Postdoctoral researcher, Dept. Biological Sciences, University of Cincinnati, USA (Dr. Nathan Morehouse) Jul 2015-Jun 2017 Postdoctoral researcher, Evolutionary Biology Centre, Uppsala University, Sweden (Drs. Frank Johansson, Anders Ödeen, & Karin Nordström) Jul 2014-Jul 2015 Visiting Professor, Dept. Ciencias Biológicas, Universidad de los Andes, Colombia Nov 2011-Dec 2013 Postdoctoral researcher, Evolutionary Biology Centre, Uppsala University, Sweden (Dr. Frank Johansson) Jun 2006-May 2010 Graduate researcher and Teaching assistant, Dept. Biología de Organismos y Sistemas, University of Oviedo, Spain (Dr. Francisco J. Ocharan) Jul 2005-Aug 2005 Intern, Servicio Regional de Investigación y Desarrollo Agroalimentario de Asturias (SERIDA), Spain (Dr. Isabel Feito Díaz) Sep 2004-Jun 2005 Undergraduate research fellow, Dept. Biología de Organismos y Sistemas, University of Oviedo, Spain (Dr. Francisco J. Ocharan) RESEARCH INTERESTS I am a behavioral ecologist, interested in the micro- and macroevolutionary processes that promote diversity. My research has explored questions on the evolution of color signals, color vision, and flight morphology. I am particularly interested in understanding the evolution of color signals, how they are perceived by intended and unintended receivers and the role of these audiences in driving population and species divergence. I also study the evolution of flight morphology because wings are large conspicuous body surfaces that can be also used as motion signal vehicles for intra- and interspecific communication. -

Critical Species of Odonata in Europe
See discussions, stats, and author profiles for this publication at: http://www.researchgate.net/publication/228966602 Critical species of Odonata in Europe ARTICLE in INTERNATIONAL JOURNAL OF ODONATOLOGY · JULY 2004 Impact Factor: 0.5 · DOI: 10.1080/13887890.2004.9748223 CITATIONS DOWNLOADS VIEWS 25 181 148 5 AUTHORS, INCLUDING: Adolfo Cordero-Rivera University of Vigo 151 PUBLICATIONS 1,594 CITATIONS SEE PROFILE Frank Suhling Technische Universität Braun… 79 PUBLICATIONS 793 CITATIONS SEE PROFILE Available from: Frank Suhling Retrieved on: 13 September 2015 Guardians of the watershed. Global status of dragonflies: critical species, threat and conservation Critical species of Odonata in Europe Göran Sahlén 1, Rafal Bernard 2, Adolfo Cordero Rivera 3, Robert Ketelaar 4 & Frank Suhling 5 1 Ecology and Environmental Science, Halmstad University, P.O. Box 823, SE-30118 Halmstad, Sweden. <[email protected]> 2 Department of General Zoology, Adam Mickiewicz University, Fredry 10, PO-61-701 Poznan, Poland. <[email protected]> 3 Departamento de Ecoloxía e Bioloxía Animal, Universidade de Vigo, EUET Forestal, Campus Universitario, ES-36005 Pontevedra, Spain. <[email protected]> 4 Dutch Butterfly Conservation. Current address: Dutch Society for the Preservation of Nature, P.O. Box 494, NL-5613 CM, Eindhoven, The Netherlands. <[email protected]> 5 Institute of Geoecology, Dpt of Environmental System Analysis, Technical University of Braunschweig, Langer Kamp 19c, D-38102 Braunschweig, Germany. <[email protected]> Key words: Odonata, dragonfly, IUCN, FFH directive, endemic species, threatened species, conservation, Europe. Abstract The status of the odonate fauna of Europe is fairly well known, but the current IUCN Red List presents only six species out of ca 130, two of which are actually out of danger today. -

The Impacts of Urbanisation on the Ecology and Evolution of Dragonflies and Damselflies (Insecta: Odonata)
The impacts of urbanisation on the ecology and evolution of dragonflies and damselflies (Insecta: Odonata) Giovanna de Jesús Villalobos Jiménez Submitted in accordance with the requirements for the degree of Doctor of Philosophy (Ph.D.) The University of Leeds School of Biology September 2017 The candidate confirms that the work submitted is her own, except where work which has formed part of jointly-authored publications has been included. The contribution of the candidate and the other authors to this work has been explicitly indicated below. The candidate confirms that appropriate credit has been given within the thesis where reference has been made to the work of others. The work in Chapter 1 of the thesis has appeared in publication as follows: Villalobos-Jiménez, G., Dunn, A.M. & Hassall, C., 2016. Dragonflies and damselflies (Odonata) in urban ecosystems: a review. Eur J Entomol, 113(1): 217–232. I was responsible for the collection and analysis of the data with advice from co- authors, and was solely responsible for the literature review, interpretation of the results, and for writing the manuscript. All co-authors provided comments on draft manuscripts. The work in Chapter 2 of the thesis has appeared in publication as follows: Villalobos-Jiménez, G. & Hassall, C., 2017. Effects of the urban heat island on the phenology of Odonata in London, UK. International Journal of Biometeorology, 61(7): 1337–1346. I was responsible for the data analysis, interpretation of results, and for writing and structuring the manuscript. Data was provided by the British Dragonfly Society (BDS). The co-author provided advice on the data analysis, and also provided comments on draft manuscripts. -

Ecological Traits of Dragonfly (Odonata) Assemblages Along An
Ann. Limnol. - Int. J. Lim. 53 (2017) 377–389 Available online at: © EDP Sciences, 2017 www.limnology-journal.org DOI: 10.1051/limn/2017019 RESEARCH ARTICLE Ecological traits of dragonfly (Odonata) assemblages along an oligotrophic Dinaric karst hydrosystem Marina Vilenica* University of Zagreb, Faculty of Teacher Education, Trg Matice hrvatske 12, 44250 Petrinja, Croatia Received: 30 March 2017; Accepted: 27 August 2017 Abstract – Ecological traits of dragonfly larvae in tufa-depositing habitats of the Dinaric karst were studied monthly over a one-year period (2007–2008). The study encompassed various lotic karst habitats (springs, mountainous rivers, streams, tufa barriers) and microhabitats (angiosperms, mosses, cobbles, sand, silt with leaf litter). The aims of the study were to identify dragonfly composition, abundance and spatial distribution, their habitat and microhabitat preferences, and to determine the most important environmental factors explaining dragonfly assemblages in the studied hydrosystem. The dragonfly fauna was composed of eight species, Onychogomphus forcipatus (Linnaeus, 1758) was the most widespread and the most numerous. Water temperature, ammonium and oxygen concentrations had the highest influence on dragonfly assemblages. The most favorable habitat type were tufa barriers, less favorable were lower lotic habitats, while dragonflies were almost completely absent from upper lotic habitats and their springs. Dragonfly larvae preferred microhabitats with inorganic substrates (i.e. cobbles and sand) and slower water velocity, while they mostly avoided mosses associated with the strongest current. This study provides an important contribution to the knowledge of dragonfly ecology in lotic habitats of the Dinaric karst. Keywords: Odonata / case study / tufa barriers / environmental factors / microhabitats freshwater habitats (Corbet, 1993; Moore, 1997; Mortimeret al., 1 Introduction 1998).Bothlarvae andadultsare generalist predatorsthat mainly feed on various small invertebrates. -

© 2016 David Paul Moskowitz ALL RIGHTS RESERVED
© 2016 David Paul Moskowitz ALL RIGHTS RESERVED THE LIFE HISTORY, BEHAVIOR AND CONSERVATION OF THE TIGER SPIKETAIL DRAGONFLY (CORDULEGASTER ERRONEA HAGEN) IN NEW JERSEY By DAVID P. MOSKOWITZ A dissertation submitted to the Graduate School-New Brunswick Rutgers, The State University of New Jersey In partial fulfillment of the requirements For the degree of Doctor of Philosophy Graduate Program in Entomology Written under the direction of Dr. Michael L. May And approved by _____________________________________ _____________________________________ _____________________________________ _____________________________________ New Brunswick, New Jersey January, 2016 ABSTRACT OF THE DISSERTATION THE LIFE HISTORY, BEHAVIOR AND CONSERVATION OF THE TIGER SPIKETAIL DRAGONFLY (CORDULEGASTER ERRONEA HAGEN) IN NEW JERSEY by DAVID PAUL MOSKOWITZ Dissertation Director: Dr. Michael L. May This dissertation explores the life history and behavior of the Tiger Spiketail dragonfly (Cordulegaster erronea Hagen) and provides recommendations for the conservation of the species. Like most species in the genus Cordulegaster and the family Cordulegastridae, the Tiger Spiketail is geographically restricted, patchily distributed with its range, and a habitat specialist in habitats susceptible to disturbance. Most Cordulegastridae species are also of conservation concern and the Tiger Spiketail is no exception. However, many aspects of the life history of the Tiger Spiketail and many other Cordulegastridae are poorly understood, complicating conservation strategies. In this dissertation, I report the results of my research on the Tiger Spiketail in New Jersey. The research to investigate life history and behavior included: larval and exuvial sampling; radio- telemetry studies; marking-resighting studies; habitat analyses; observations of ovipositing females and patrolling males, and the presentation of models and insects to patrolling males. -

Indiana County Natural Heritage Inventory
Indiana County Natural Heritage Inventory Update 2021 Indiana County Natural Heritage Inventory 2021 Update Anna Johnson and Christopher Tracey, editors Prepared for: Southwest Pennsylvania Commission 112 Washington Pl #500 Pittsburgh, PA 15219 Prepared by: Pennsylvania Natural Heritage Program 800 Waterfront Drive Pittsburgh, PA 15222 Please cite this Natural Heritage Inventory report as: Johnson, Anna and Christopher Tracey, editors. 2021. Indiana County Natural Heritage Inventory. Pennsylvania Natural Heritage Program. Pittsburgh, PA. 1 ACKNOWLEDGEMENTS We would like to acknowledge the citizens and landowners of Indiana County and surrounding areas who volunteered infor- mation, time, and effort to the inventory and granted permission to access land. A big thank you goes to those who suggested areas of interest, provided data, and assisted with field surveys. Additional thanks goes to Ryan Gordon of the Southwest Pennsylvania Commission for providing support for this project. Advisory Committee to the 2021 update to the Indiana County Natural Heritage Inventory: • Byron Stauffer Jr. — Indiana County Executive Director of Planning and Development • Jeff Raykes — Indiana County Planner • Josh Krug — Indiana County Planner We want to recognize the Pennsylvania Natural Heritage Program and NatureServe for providing the foundation for the work that we perform for these studies. Current and former PNHP staff that contributed to this report includes JoAnn Albert, Jaci Braund, Charlie Eichelberger, Kierstin Carlson, Mary Ann Furedi, Steve Grund, Amy Jewitt, Anna Johnson, Susan Klugman, John Kunsman, Betsy Leppo, Jessica McPherson, Molly Moore, Ryan Miller, Greg Podniesinski, Megan Pulver, Erika Schoen, Scott Schuette, Emily Szoszorek, Kent Taylor, Christopher Tracey, Natalie Virbitsky, Jeff Wagner, Denise Watts, Joe Wisgo, Pete Woods, David Yeany, and Ephraim Zimmerman. -

Anisoptera: Cordulegastridae)
Odonatologica 30(4): 395-409 December 1, 2001 Larval habitats and longitudinaldistribution patterns of Cordulegasterheros Theischinger and C. bidentata Sélys in an Austrian forest stream (Anisoptera: Cordulegastridae) C. Lang,H. Müller and J.A. Waringer Institute of Ecology and Nature Conservation, University of Vienna, Althanstraße 14, A-1090 Vienna, Austria Received September 10, 2000 /Revised and Accepted March 5, 2001 From May 1997 to April 1998 larvae were recorded at the Weidlingbach,a fourth order 12 tributary ofthe Danube nr Vienna, at sampling stations from source to mouth. From the 14 larval instars for the 5 based head could be identified in reported genus, (F to F-4; on width) both head larvae than in — Both spp.; widths of $ were significantlylarger 3 . spp. were most abundantwithin sand sedimentswith size of2.04 in C. bidentata medium a meangrain (QJ mm and 2.79 mm in C. heros. Mean water depths and nose current speeds measured at larval microhabitats were 4.4 cm and 2.3cm s’ 1 ((C. bidentata) and 5.6 cm and 2.6cm s' 1 ((C. heros). During the winter months larvae chose the water depths slightly deeper than during summer. Throughoutthe observation period, a high proportion ofthe larvae (C. bidentata; 70-100%; C. hems'. 41-90%) were burrowed in sandy sediments,either totally or displaying the typical ambush posture with only head and anal pyramid visible. In winter,theproportion ofburrowing larvae than in — C. larvae abundant was insignificantly higher summer. bidentata weremost near the sources, preferringfirst order stream sections (discharge0.1-3.2 Is 1 )with highhardness (up to 34 German degrees) and conductivity (up to 1100 pS cm 1 ) and a high proportion of fine C. -

Odonata: Cordulegaster Leach, 1815) V Slovenskej Republike
PRÍSPEVOK K BIONÓMII PÁSIKAVCOV (ODONATA: CORDULEGASTER LEACH, 1815) V SLOVENSKEJ REPUBLIKE Stanislav DAVID1, Kornélia PETROVIČOVÁ2 1Ústav krajinnej ekológie SAV, pobočka Nitra, Akademická 2, 949 10 Nitra e-mail: [email protected] 2Katedra environmentalistiky a zoológie, Fakulta agrobiológie a potravinových zdrojov, Slovenská poľnohospodárska univerzita, Tr. A. Hlinku 2, SK-949 76 e-mail: [email protected] Abstract: A golden-ringed dragonfly (Odonata, Cordulegastridae) is represented by two species Cordulegaster bidentata and C. heros in the territory of Slovakia. As for both species, we evaluated hypsometry of their occurrence, the habitat preferences and the structure of the odonatocenoses. The dataset for C. bidentata species consists of 103 sites and C. heros species is from 124 localities. C. bidentata is evenly spread in the mountainous areas of Slovakia. C. heros has had the discontinuous occurrence in the Male Karpaty Mts. and Slovenské rudohorie Mts. The average altitude of localities of the C. bidentata is 563.3 m a. s. l., nevertheless for the C. heros it is 304.5 m. The significant habitat factors of examined species (p = 0.05) are altitude, the forest margin, the fens with watersurface, epirithron and hyporithron respectively. The golden-ringed dragonfly has been examined together with other 15 species, but only on peat marshes there were 13 nonreproduced ones. The Cordulegaster heros larvae occur together only with C. bidentata and Calopteryx virgo larvae. Our results proved the published habitat preferences of C. bidentata and C. heros in Slovakia. Key words: Cordulegaster species, distribution, habitats, evaluation, Slovakia Úvod Rod pásikavce (Cordulegaster Leach, 1815) obsahuje 29 druhov (Schoor, Paulson, 2018) s holoarktickým rozšírením. -
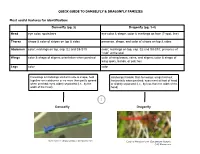
Dragonfly (Pg. 3-4) Head Eye Color
QUICK GUIDE TO DAMSELFLY & DRAGONFLY FAMILIES Most useful features for identification: Damselfly (pg. 2) Dragonfly (pg. 3-4) Head eye color; spots/bars eye color & shape; color & markings on face (T-spot, line) Thorax shape & color of stripes on top & sides presence, shape, and color of stripes on top & sides Abdomen color; markings on top, esp. S2 and S8-S10 color; markings on top, esp. S2 and S8-S10; presence of “club” at the end Wings color & shape of stigma; orientation when perched color of wing bases, veins, and stigma; color & shape of wing spots, bands, or patches Legs color color Forewings & hindwings similar in size & shape, held Hindwings broader than forewings; wings held out together over abdomen or no more than partly spread horizontally when perched; eyes meet at front of head when perched; eyes widely separated (i.e., by the or slightly separated (i.e., by less than the width of the width of the head) head) 1 Damselfly Dragonfly Vivid Dancer (Argia vivida); CAS Mazzacano Cardinal Meadowhawk (Sympetrum illotum); CAS Mazzacano DAMSELFLIES Wings narrow, stalked at base 2 Wings broad, colored, not stalked at base 3 Wings held askew Wings held together Broad-winged Damselfly when perched when perched (Calopterygidae); streams Spreadwing Pond Damsels (Coenagrionidae); (Lestidae); ponds ponds, streams Dancer (Argia); Wings held above abdomen; vivid colors streams 4 Wings held along Bluet (Enallagma); River Jewelwing (Calopteryx aequabilis); abdomen; mostly blue ponds CAS Mazzacano Dark abdomen with blue Forktail (Ischnura); tip; -

Check List of First Recorded Dragonfly (Odonata: Anisoptera) Fauna of District Lower Dir, Khyber Pakhtunkhwa, Pakistan
Arthropods, 2014, 3(2): 120-126 Article Check list of first recorded dragonfly (Odonata: Anisoptera) fauna of District Lower Dir, Khyber Pakhtunkhwa, Pakistan Farzana Perveen1, Anzela Khan2, Sayed Abdul Rauf3 1Departments of Zoology, Shaheed Benazir Bhutto University (SBBU), Main Campus, Sheringal, Khyber Pakhtunkhwa, Pakistan 2Beaconhouse School System, Margalla Campus (BMI-G), H-8, Islamabad, Pakistan 3Departments of Zoology, Shaheed Benazir Bhutto University (SBBU), Main Campus, Sheringal, Khyber Pakhtunkhwa, Pakistan E-mail: [email protected] Received 5 March 2014; Accepted 10 April 2014; Published online 1 June 2014 Abstract The dragonflies (Odonata: Anisoptera) are large, intermediate to small size, having different colours and variable morphological characters. They also carry ornamental and environmental indicator values. The first recorded, the collection of 318 dragonflies was made during May-July 2011 from district Lower Dir, Khyber Pakhtunkhwa, Pakistan. Among them 11 species of dragonflies were identified belonging to 3 families. The golden-ringed, Cordulegaster brevistigma brevistigma Selys is belonging to family Cordulegasteridae and Clubtails, Onychogomphus bistrigatus Selys is belonging to family Gomophidaed. The spine-legged redbolt, Rhodothemis rufa (Rambur); black-tailed skimmer, Orthetrum cancellatum Linnaeus; blue or black-percher, Diplacodes lefebvrei (Ramber); ground-skimmer, Diplacodes trivialis Rambur; common red-skimmer, Orthetrum pruinosum neglectum (Rambur); triangle-skimmer, Orthetrum triangulare triangulare -

Journal Vol 26 No 2, October 2010
J. Br. Dragonfly Society, Volume 26 No. 2, October 2010 Journal of the CONTENTS DAVID CHELMICK - Studying British dragonflies in the British Dragonfly Society 1970s: the wilderness years .............................................. 57 Volume 26 Number 2 October 2010 BARRY NATTRESS - Folding wing behaviour in Cordulagaster boltonii (Donovan) ............................................................. 64 DAVID CHELMICK - Species Review 4: The Scarce Emerald Damselfly Lestes dryas Kirby with notes on the family Lestidae in the Western Palearctic ....................................66 JONATHAN. R. DIXON & DOROTHY E. GENNARD - The influence of meteorological conditions on the flight activity of the Blue-tailed Damselfly Ischnura elegans (Vander Linden), the Azure Damselfly Coenagrion puella (Linnaeus) and the Emerald Damselfly Lestes sponsa (Hansemann) ..... .............................................................................................. 83 ADRIAN J. PARR -. Migrant and dispersive dragonflies in Britain during 2009 ............................................................97 PAM TAYLOR & DAVE SMALLSHIRE - A change in status of the Dainty Damselfly Coenagrion scitulum (Rambur) in the United Kingdom ………....................................................107 Corrigendum ..........................................................................i The aims of the British Dragonfly Society (BDS) are to promote and encourage the study and conservation INSTRUCTIONS TO AUTHORS of Odonata and their natural habitats, especially in the -

Damselflies & Dragonflies of the Cairngorms
Damselflies & Dragonflies of the Cairngorms Male and female Northern damselfly in tandem An identification guide Dragonflies are amazing insects that combine stunning colours with awesome aerial displays. This guide will help you identify the 13 species of dragonfly and damselfly found in and around the Cairngorms National Park. You can get involved and record all these species and help put together a national atlas of dragonflies. Published by The Cairngorms LBAP 1M 2M 3M 1F 2F 3F Damselflies Delicate 2. Large Red Damselfly vegetation usually below the insects, weak fluttery flight. Tot: 33 – 36mm tops of plants, and avoid large Eyes on each side of oblong Range: Cairngorms-wide areas of open water. The head, wings usually held Rarity: common underside of the eyes and closed at rest. Habitat: still or slow face are bright green. Males 1. Emerald Damselfly moving water have two short black lines on Total body length (Tot): Seen: May to August each side of the thorax, and 35 – 39mm A distinctive red and black a spear-shaped mark and 2 Range: Cairngorms-wide damselfly that is often the black lines on the second Rarity: common first to be seen in late spring. abdominal segment. Females Habitat: well vegetated Females are darker with are pea-green with the 2 standing water, ditches and black and yellow bands on thorax lines. loch edges the abdomen. Males defend 4. Common Blue Seen: June to September their territories vigorously. Damselfly A slender species, with a 3. Northern Damselfly Tot: 29 – 36mm weak, fluttery flight. Males Tot: 31 – 33mm Range: Cairngorms-wide are metallic green with blue Range: limited to only 30 Rarity: common segments 9 and 10 on the sites in the Cairngorms Habitat: most wetlands rear of the abdomen.