G-CSF Protects Motoneurons Against Axotomy-Induced Apoptotic Death In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Therapeutic Class Overview Colony Stimulating Factors
Therapeutic Class Overview Colony Stimulating Factors Therapeutic Class Overview/Summary: This review will focus on the granulocyte colony stimulating factors (G-CSFs) and granulocyte- macrophage colony stimulating factors (GM-CSFs).1-5 Colony-stimulating factors (CSFs) fall under the naturally occurring glycoprotein cytokines, one of the main groups of immunomodulators.6 In general, these proteins are vital to the proliferation and differentiation of hematopoietic progenitor cells.6-8 The G- CSFs commercially available in the United States include pegfilgrastim (Neulasta®), filgrastim (Neupogen®), filgrastim-sndz (Zarxio®), and tbo-filgrastim (Granix®). While filgrastim-sndz and tbo- filgrastim are the same recombinant human G-CSF as filgrastim, only filgrastim-sndz is considered a biosimilar drug as it was approved through the biosimilar pathway. At the time tbo-filgrastim was approved, a regulatory pathway for biosimilar drugs had not yet been established in the United States and tbo-filgrastim was filed under its own Biologic License Application.9 Only one GM-CSF is currently available, sargramostim (Leukine). These agents are Food and Drug Administration (FDA)-approved for a variety of conditions relating to neutropenia or for the collection of hematopoietic progenitor cells for collection by leukapheresis.1-5 Due to the pathway taken, tbo-filgrastim does not share all of the same indications as filgrastim and these two products are not interchangeable. It is important to note that although filgrastim-sndz is a biosimilar product, and it was approved with all the same indications as filgrastim at the time, filgrastim has since received FDA-approval for an additional indication that filgrastim-sndz does not have, to increase survival in patients with acute exposure to myelosuppressive doses of radiation.1-3A complete list of indications for each agent can be found in Table 1. -

Comparison Between Filgrastim and Lenograstim Plus Chemotherapy For
Bone Marrow Transplantation (2010) 45, 277–281 & 2010 Macmillan Publishers Limited All rights reserved 0268-3369/10 $32.00 www.nature.com/bmt ORIGINAL ARTICLE Comparison between filgrastim and lenograstim plus chemotherapy for mobilization of PBPCs R Ria, T Gasparre, G Mangialardi, A Bruno, G Iodice, A Vacca and F Dammacco Department of Biomedical Sciences and Human Oncology, Section of Internal Medicine and Clinical Oncology, University of Bari Medical School, Bari, Italy Recombinant human (rHu) G-CSF has been widely used during transplantation.4 However, the use of G-CSF after to treat neutropenia and mobilize PBPCs for their autologous PBPC transplantation has been queried, as its autologous and allogeneic transplantation. It shortens further reduction in time to a safe neutrophil count5,6 does neutropenia and thus reduces the frequency of neutropenic not always imply fewer significant clinical events, such as fever. We compared the efficiency of glycosylated rHu infections, length of hospitalization, extrahematological and non-glycosylated Hu G-CSF in mobilizing hemato- toxicities or mortality.7,8 Even so, the ASCO guidelines still poietic progenitor cells (HPCs). In total, 86 patients were recommend the use of growth factors after autologous consecutively enrolled for mobilization with CY plus either PBPC transplantation.9 glycosylated or non-glycosylated G-CSF, and under- G-CSF induces the proliferation and differentiation of went leukapheresis. The HPC content of each collection, myeloid precursor cells, and also provides a functional toxicity, days of leukapheresis needed to reach the activity that influences chemotaxis, respiratory burst and minimum HPC target and days to recover WBC (X500 Ag expression of neutrophils. -

©Ferrata Storti Foundation
Stem Cell Transplantation • Research Paper Single-dose pegfilgrastim for the mobilization of allogeneic CD34+ peripheral blood progenitor cells in healthy family and unrelated donors Frank Kroschinsky Background and Objectives. Short-term treatment with granulocyte colony-stimulating Kristina Hölig factor (G-CSF) has been established as the standard regimen for mobilizing allogeneic Kirsten Poppe-Thiede peripheral blood progenitor cells (PBPC) from healthy donors. The pegylated form of fil- Kristin Zimmer grastim (pegfilgrastim) has a longer elimination half-life because of decreased serum Rainer Ordemann clearance and might be a convenient alternative for stem cell mobilization. Matthias Blechschmidt Uta Oelschlaegel Design and Methods. Twenty-five family (n=15) or unrelated (n=10) healthy donors received a single-dose of 12 mg pegfilgrastim for mobilization of allogeneic PBPC. Martin Bornhauser + Gabi Rall Donors with inadequate mobilization (blood CD34 cells ≤5/µL on day 3 or ≤20/µL on Claudia Rutt day 4) were given additional daily doses of 10 µg/kg conventional filgrastim. Gerhard Ehninger Leukapheresis was planned to start on day 5. Results. All harvests were completed successfully. In 20 out of 25 donors (80 %) only a single apheresis was necessary. Additional non-pegylated filgrastim had to be given to only one 74-year old family donor. The maximum concentration of circulating CD34+ + cells occurred on day 5 (median 67/µL, range 10-385/µL). The median yield of CD34 cells was 9.3 (range 3.2-39.1) 106/kg of the recipient´s body weight. The median × 8 number of T cells in the apheresis products was 3.9 (range 2.7-10.8)×10 /kg. -
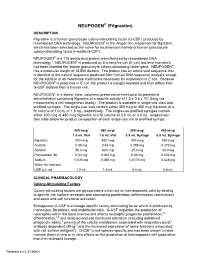
NEUPOGEN® (Filgrastim) DESCRIPTION Filgrastim Is a Human Granulocyte Colony-Stimulating Factor (G-CSF)‚ Produced by Recombinant DNA Technology
NEUPOGEN® (Filgrastim) DESCRIPTION Filgrastim is a human granulocyte colony-stimulating factor (G-CSF)‚ produced by recombinant DNA technology. NEUPOGEN® is the Amgen Inc. trademark for filgrastim‚ which has been selected as the name for recombinant methionyl human granulocyte colony-stimulating factor (r-metHuG-CSF). NEUPOGEN® is a 175 amino acid protein manufactured by recombinant DNA technology.1 NEUPOGEN® is produced by Escherichia coli (E coli) bacteria into which has been inserted the human granulocyte colony-stimulating factor gene. NEUPOGEN® has a molecular weight of 18‚800 daltons. The protein has an amino acid sequence that is identical to the natural sequence predicted from human DNA sequence analysis‚ except for the addition of an N-terminal methionine necessary for expression in E coli. Because NEUPOGEN® is produced in E coli‚ the product is nonglycosylated and thus differs from G-CSF isolated from a human cell. NEUPOGEN is a sterile‚ clear‚ colorless‚ preservative-free liquid for parenteral administration containing filgrastim at a specific activity of 1.0 ± 0.6 x 108 U/mg (as measured by a cell mitogenesis assay). The product is available in single-use vials and prefilled syringes. The single-use vials contain either 300 mcg or 480 mcg filgrastim at a fill volume of 1.0 mL or 1.6 mL, respectively. The single-use prefilled syringes contain either 300 mcg or 480 mcg filgrastim at a fill volume of 0.5 mL or 0.8 mL, respectively. See table below for product composition of each single-use vial or prefilled syringe. 300 mcg/ 480 mcg/ 300 mcg/ 480 mcg/ 1.0 mL Vial 1.6 mL Vial 0.5 mL Syringe 0.8 mL Syringe filgrastim 300 mcg 480 mcg 300 mcg 480 mcg Acetate 0.59 mg 0.94 mg 0.295 mg 0.472 mg Sorbitol 50.0 mg 80.0 mg 25.0 mg 40.0 mg Polysorbate 80 0.04 mg 0.064 mg 0.02 mg 0.032 mg Sodium 0.035 mg 0.056 mg 0.0175 mg 0.028 mg Water for Injection USP q.s. -

WO 2018/218208 Al 29 November 2018 (29.11.2018) W !P O PCT
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date WO 2018/218208 Al 29 November 2018 (29.11.2018) W !P O PCT (51) International Patent Classification: Fabio; c/o Bruin Biosciences, Inc., 10225 Barnes Canyon A61K 31/00 (2006.01) A61K 47/00 (2006.01) Road, Suite A104, San Diego, California 92121-2734 (US). BEATON, Graham; c/o Bruin Biosciences, Inc., 10225 (21) International Application Number: Barnes Canyon Road, Suite A104, San Diego, California PCT/US20 18/034744 92121-2734 (US). RAVULA, Satheesh; c/o Bruin Bio (22) International Filing Date: sciences, Inc., 10225 Barnes Canyon Road, Suite A l 04, San 25 May 2018 (25.05.2018) Diego, Kansas 92121-2734 (US). (25) Filing Language: English (74) Agent: MALLON, Joseph J.; 2040 Main Street, Four teenth Floor, Irvine, California 92614 (US). (26) Publication Langi English (81) Designated States (unless otherwise indicated, for every (30) Priority Data: kind of national protection available): AE, AG, AL, AM, 62/5 11,895 26 May 2017 (26.05.2017) US AO, AT, AU, AZ, BA, BB, BG, BH, BN, BR, BW, BY, BZ, 62/5 11,898 26 May 2017 (26.05.2017) US CA, CH, CL, CN, CO, CR, CU, CZ, DE, DJ, DK, DM, DO, (71) Applicants: BRUIN BIOSCIENCES, INC. [US/US]; DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, HN, 10225 Barnes Canyon Road, Suite A104, San Diego, HR, HU, ID, IL, IN, IR, IS, JO, JP, KE, KG, KH, KN, KP, California 92121-2734 (US). -

Comparative Safety of Filgrastim Versus Sargramostim in Patients Receiving Myelosuppressive Chemotherapy
Comparative Safety of Filgrastim versus Sargramostim in Patients Receiving Myelosuppressive Chemotherapy Gary Milkovich, B.S., Ronald J. Moleski, Pharm.D., John F. Reitan, Pharm.D., David M. Dunning, M.D., Gene A. Gibson, Pharm.D., Thomas A. Paivanas, M.H.S.A., Susan Wyant, Pharm.D., and R. Jake Jacobs, M.P.A. Study Objective. To compare rates of adverse events with filgrastim versus sargramostim when given prophylactically to patients receiving myelosuppressive chemotherapy. Design. Retrospective review with center crossover. Setting. Ten United States outpatient chemotherapy centers. Patients. Four hundred ninety patients treated for lung, breast, lymphatic system, or ovarian tumors. Intervention. Prophylactic use of filgrastim or sargramostim, with dosages at investigator discretion. Measurements and Main Results. The frequency and severity of adverse events and the frequency of switching to the alternative CSF were assessed. There was no difference in infectious fever. Fever unexplained by infection was more common with sargramostim (7% vs 1%, p<0.001), as were fatigue, diarrhea, injection site reactions, other dermatologic disorders, and edema (all p<0.05). Skeletal pain was more frequent with filgrastim (p=0.06). Patients treated with sargramostim switched to the alternative agent more often (p<0.001). Conclusion. Adverse events were less frequent with filgrastim than with sargramostim, suggesting that quality of life and treatment costs also may differ. (Pharmacotherapy 2000;20(12):1432–1440) Colony-stimulating factors (CSFs) are therapy. Bone pain is the most commonly recommended in certain situations to reduce the reported adverse event with filgrastim,6, 7 but likelihood of febrile neutropenia induced by exacerbation of preexisting inflammatory myelosuppressive chemotherapy.1 Recombinant conditions,8–10 rash,11 allergic reactions,12 Sweet forms of granulocyte (G)-CSF and granulocyte- syndrome,13 and injection site reactions14 have macrophage (GM)-CSF are synthesized using a been reported. -

NEUPOGEN (Filgrastim) Patient Guide
NEUPOGEN® (filgrastim) Patient Guide What is NEUPOGEN®? NEUPOGEN® should be injected at least 24 hours If you are giving someone else NEUPOGEN® NEUPOGEN® is a man-made form of granulocyte before your next dose of chemotherapy. injections, it is important that you know how to inject colony-stimulating factor (G-CSF), which is made There is more information about NEUPOGEN® in the NEUPOGEN®, how much to inject, and how often to using the bacteria Escherichia coli. G-CSF is a substance Physician Package Insert. If you have any questions, you inject NEUPOGEN®. naturally produced by the body. It stimulates the growth should talk to your doctor. NEUPOGEN® is available as a liquid in vials of neutrophils (nu-tro-fils), a type of white blood cell What are possible serious side effects of or in prefilled syringes. When you receive your important in the body’s fight against infection. NEUPOGEN®? NEUPOGEN®, always check to see that: ® What is NEUPOGEN used for? • Spleen Rupture. Your spleen may become enlarged • The name NEUPOGEN® appears on the package NEUPOGEN® is a prescription medication used to and can rupture while taking NEUPOGEN®. A and vial or prefilled syringe label. reduce the risk of infection in patients with some ruptured spleen can cause death. The spleen is located • The expiration date on the vial or prefilled syringe tumors, who are receiving strong chemotherapy that in the upper left section of your stomach area. Call label has not passed. You should not use a vial or may cause severe neutropenia (nu-tro-peen-ee-ah), your doctor right away if you/your child has pain in prefilled syringe after the date on the label. -

Granulocyte Colony-Stimulating Factor (G-CSF)
British Journal of Cancer (1999) 80(1/2), 229–235 © 1999 Cancer Research Campaign Article no. bjoc.1998.0344 Granulocyte colony-stimulating factor (G-CSF) transiently suppresses mitogen-stimulated T-cell proliferative response E Reyes1, I García-Castro2, F Esquivel1, J Hornedo2, H Cortes-Funes2, J Solovera3 and M Alvarez-Mon1 1Medicine/Immune System Diseases Oncology Service, Department of Medicine ‘Principe de Asturias’ University Hospital, Alcalá University, Carretera Madrid- Barcelona, Km 33.600, 28871 Alcalá de Henares, Madrid, Spain; 2Medical Oncology Department, ‘12 de Octubre’ University Hospital, Madrid, Spain; 3Amgen, S.A. Barcelona, Spain Summary Granulocyte colony-stimulation factor (G-CSF) is a cytokine that selectively promotes growth and maturation of neutrophils and may modulate the cytokine response to inflammatory stimuli. The purpose of this study was to examine the effect of G-CSF on ex vivo peripheral blood mononuclear cell (PBMC) functions. Ten patients with breast cancer were included in a clinical trial in which r-metHuG-CSF was administrered daily for 5 days to mobilize peripheral blood stem cells. Ten healthy women were also included as controls. Our data show that G-CSF treatment induces an increase in peripheral blood leucocyte, neutrophil, lymphocyte and monocyte counts. We have found a modulation in the percentages of CD19+, CD45+CD14+, CD4+CD45RA+ and CD4+CD45RO+ cells in PBMC fractions during G-CSF treatment. We have also found a significant reduction in the proliferative response of PBMC to mitogenic stimulation that reverted 14 days after the fifth and the last dose of G-CSF. Furthermore, it was not associated with significant changes in the pattern of cytokine production. -

INN Working Document 05.179 Update December 2010
INN Working Document 05.179 Update December 2010 International Nonproprietary Names (INN) for biological and biotechnological substances (a review) INN Working Document 05.179 Distr.: GENERAL ENGLISH ONLY 12/2010 International Nonproprietary Names (INN) for biological and biotechnological substances (a review) Programme on International Nonproprietary Names (INN) Quality Assurance and Safety: Medicines Essential Medicines and Pharmaceutical Policies (EMP) International Nonproprietary Names (INN) for biological and biotechnological substances (a review) © World Health Organization 2010 All rights reserved. Publications of the World Health Organization can be obtained from WHO Press, World Health Organization, 20 Avenue Appia, 1211 Geneva 27, Switzerland (tel.: +41 22 791 3264; fax: +41 22 791 4857; e-mail: [email protected]). Requests for permission to reproduce or translate WHO publications – whether for sale or for noncommercial distribution – should be addressed to WHO Press, at the above address (fax: +41 22 791 4806; e-mail: [email protected]). The designations employed and the presentation of the material in this publication do not imply the expression of any opinion whatsoever on the part of the World Health Organization concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Dotted lines on maps represent approximate border lines for which there may not yet be full agreement. The mention of specific companies or of certain manufacturers’ products does not imply that they are endorsed or recommended by the World Health Organization in preference to others of a similar nature that are not mentioned. Errors and omissions excepted, the names of proprietary products are distinguished by initial capital letters. -

And G-CSF (filgrastim) After Cyclophosphamide (4 G/M2) Enhance the Peripheral Blood Progenitor Cell Harvest: Results of Two Randomized Studies Including 120 Patients
Bone Marrow Transplantation (2006) 38, 275–284 & 2006 Nature Publishing Group All rights reserved 0268-3369/06 $30.00 www.nature.com/bmt ORIGINAL ARTICLE Low doses of GM-CSF (molgramostim) and G-CSF (filgrastim) after cyclophosphamide (4 g/m2) enhance the peripheral blood progenitor cell harvest: results of two randomized studies including 120 patients P Quittet1, P Ceballos1, E Lopez1,ZYLu2, P Latry1, C Becht1, E Legouffe1, N Fegueux1, C Exbrayat1, DPouessel 1, V Rouille´ 1, JP Daures3, B Klein2,4 and JF Rossi1,4 1Service He´matologie et Oncologie me´dicale, Hopital Lapeyronie, Montpellier, France; 2Unite´ de The´rapie Cellulaire, Hopital St-Eloi, Montpellier, France; 3Unite´ de Biostatistiques, IURC, Montpellier, France and 4INSERM U475, Montpellier, France The use of a combination of G-CSF and GM-CSF versus of hematopoietic growth factors (HGFs) alone or in G-CSF alone, after cyclophosphamide (4 g/m2) was combination with chemotherapy, in which case a higher compared in two randomized phase III studies, including yield of CD34 þ cells can be reached and collected.6 It is 120 patients. In study A, 60 patients received 5 Â 2 lg/kg/ generally recognized that granulocyte colony-stimulating day of G-CSF and GM-CSF compared to 5 lg/kg/day of factor (G-CSF) as a single agent mobilizes more CD34 þ G-CSF. In study B, 60 patients received 2.5 Â 2 lg/kg/ cells than does granulocyte–macrophage colony stimulating day G-CSF and GM-CSF compared to G-CSF alone factor (GM-CSF).7–9 Other HGFs, such as interleukin-3,10 (5 lg/kg/day). -

Lonquex; INN-Lipegfilgrastim
EMA/236686/2019 EMEA/H/C/002556 Lonquex (lipegfilgrastim) An overview of Lonquex and why it is authorised in the EU What is Lonquex and what is it used for? Lonquex is a medicine that contains the active substance lipegfilgrastim. It is used to reduce the duration of neutropenia (low levels of neutrophils, a type of white blood cell) and the occurrence of febrile neutropenia (neutropenia with fever) in adult cancer patients receiving cytotoxic chemotherapy. Cytotoxic chemotherapy (medicines that kill fast growing cells) commonly causes neutropenia because in addition to cancer cells, it also kills other fast-growing cells such as neutrophils, leaving the patient at risk of infections. Lonquex is not used in patients receiving chemotherapy for chronic myeloid leukaemia (a cancer of the white blood cells) and myelodysplastic syndromes (a disease that can develop into a leukaemia). How is Lonquex used? Lonquex is available as an injection in pre-filled syringes. The injections should be given under the skin in the abdomen, upper arm or thigh. One 6-mg dose is given in each chemotherapy cycle around 24 hours after chemotherapy. The medicine can only be obtained with a prescription and treatment should be started and supervised by a doctor experienced in treating cancer and blood disorders. Patients may be able to inject the medicine themselves once they have been properly trained but the first injection should be under the direct supervision of a doctor. For more information, see the package leaflet or contact your doctor or pharmacist. How does Lonquex work? The active substance in Lonquex, lipegfilgrastim, is similar to granulocyte colony stimulating factor (G‑ CSF), a naturally occurring protein in the body that stimulates the production of white blood cells including neutrophils in the bone marrow. -

NEUPOGEN (Filgrastim) Is a 175 Amino Acid Human Granulocyte Colony-Stimulating Factor (G-CSF) Manufactured by Recombinant DNA Technology
HIGHLIGHTS OF PRESCRIBING INFORMATION ---------------------DOSAGE FORMS AND STRENGTHS--------------------- These highlights do not include all the information needed to use Vial NEUPOGEN safely and effectively. See full prescribing information for • Injection: 300 mcg/mL in a single-use vial (3) NEUPOGEN. • Injection: 480 mcg/1.6 mL in a single-use vial (3) NEUPOGEN® (filgrastim) injection, for subcutaneous or intravenous use Prefilled Syringe Initial U.S. Approval: 1991 • Injection: 300 mcg/0.5 mL in a single-use prefilled syringe (3) • Injection: 480 mcg/0.8 mL in a single-use prefilled syringe (3) ---------------------------RECENT MAJOR CHANGES-------------------------- Indications and Usage (1.6) 03/2015 -------------------------------CONTRAINDICATIONS----------------------------- Dosage and Administration (2.5) 03/2015 Patients with a history of serious allergic reactions to human granulocyte colony-stimulating factors such as filgrastim or pegfilgrastim. (4) --------------------------- INDICATIONS AND USAGE--------------------------- NEUPOGEN is a leukocyte growth factor indicated to ---------------------------WARNINGS AND PRECAUTIONS------------------- • Decrease the incidence of infection‚ as manifested by febrile neutropenia‚ • Fatal splenic rupture: Evaluate patients who report left upper abdominal or in patients with nonmyeloid malignancies receiving myelosuppressive anti shoulder pain for an enlarged spleen or splenic rupture. (5.1) cancer drugs associated with a significant incidence of severe neutropenia • Acute respiratory distress syndrome (ARDS): Evaluate patients who with fever (1.1) develop fever and lung infiltrates or respiratory distress for ARDS. • Reduce the time to neutrophil recovery and the duration of fever, following Discontinue NEUPOGEN in patients with ARDS. (5.2) induction or consolidation chemotherapy treatment of patients with acute • Serious allergic reactions, including anaphylaxis: Permanently discontinue myeloid leukemia (AML) (1.2) NEUPOGEN in patients with serious allergic reactions.