VEGETATION and MARINE ECOSYSTEM CHANGE Early Pleistocene in the Netherlands
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Atlantic Constrain Laurentide Ice Sheet Extent During Marine Isotope Stage 3
Sea-level records from the U.S. mid- Atlantic constrain Laurentide Ice Sheet extent during Marine Isotope Stage 3 The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Pico, T, J. R. Creveling, and J. X. Mitrovica. 2017. “Sea-level records from the U.S. mid-Atlantic constrain Laurentide Ice Sheet extent during Marine Isotope Stage 3.” Nature Communications 8 (1): 15612. doi:10.1038/ncomms15612. http://dx.doi.org/10.1038/ ncomms15612. Published Version doi:10.1038/ncomms15612 Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:33490941 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA ARTICLE Received 7 Dec 2016 | Accepted 11 Apr 2017 | Published 30 May 2017 DOI: 10.1038/ncomms15612 OPEN Sea-level records from the U.S. mid-Atlantic constrain Laurentide Ice Sheet extent during Marine Isotope Stage 3 T. Pico1, J.R. Creveling2 & J.X. Mitrovica1 The U.S. mid-Atlantic sea-level record is sensitive to the history of the Laurentide Ice Sheet as the coastline lies along the ice sheet’s peripheral bulge. However, paleo sea-level markers on the present-day shoreline of Virginia and North Carolina dated to Marine Isotope Stage (MIS) 3, from 50 to 35 ka, are surprisingly high for this glacial interval, and remain unexplained by previous models of ice age adjustment or other local (for example, tectonic) effects. -

Provenance of Early Pleistocene Sediments Based on a High-Resolution Sedimentological Dataset of Borehole Petten, Southern North Sea
Provenance of Early Pleistocene sediments based on a high-resolution sedimentological dataset of borehole Petten, southern North Sea Hao Ding 6559972 A thesis submitted for the degree of MSc Earth, Life and Climate Utrecht University July, 2020 1 Preface and Acknowledgements This thesis was written as part of the Master of Science degree in Earth Sciences, program Earth, Life and Climate, at Utrecht University. The research was performed under the supervision of Kim Cohen and Wim Hoek, also with help of Timme Donders and Alexander Houben. This research, together with a parallel research conducted by Lissane Krom, contributes to a larger project in paleoenvironment reconstruction of the Early Pleistocene in the southern North Sea Basin, based on the Petten Borehole 1. This thesis focused on sedimentology, and the parallel thesis by Krom focused on palynology. For the process of this project, I would like to express my gratitude to my supervisors Kim Cohen and Wim Hoek for their great effort on the research and lab work guidance. All the redactional and scientific comments on my writing from Kim Cohen are highly appreciated. I would also like to thank Timme Donders, Alexander Houben and Lisanne Krom for all the cross-disciplinary discussions as well as the assistance on my final presentation. Last but not least, I would like to thank my family and friends, who have been supporting me as always during this Covid-19 pandemic. Studying abroad has been tough, but I really appreciate all the love and encouragement around me, no matter how far apart we are. 2 Abstract In 2018, a fairly complete core (Petten BH 1) reaching over 300 meters into the unconsolidated, dominantly sandy Pleistocene sequence was recovered in the northwest of the Netherlands, in the coastal dune area along the present North Sea. -

The Easternmost Occurrence of Mammut Pacificus (Proboscidea: Mammutidae), Based on a Partial Skull from Eastern Montana, USA
The easternmost occurrence of Mammut pacificus (Proboscidea: Mammutidae), based on a partial skull from eastern Montana, USA Andrew T. McDonald1, Amy L. Atwater2, Alton C. Dooley Jr1 and Charlotte J.H. Hohman2,3 1 Western Science Center, Hemet, CA, United States of America 2 Museum of the Rockies, Montana State University, Bozeman, MT, United States of America 3 Department of Earth Sciences, Montana State University, Bozeman, MT, United States of America ABSTRACT Mammut pacificus is a recently described species of mastodon from the Pleistocene of California and Idaho. We report the easternmost occurrence of this taxon based upon the palate with right and left M3 of an adult male from the Irvingtonian of eastern Montana. The undamaged right M3 exhibits the extreme narrowness that characterizes M. pacificus rather than M. americanum. The Montana specimen dates to an interglacial interval between pre-Illinoian and Illinoian glaciation, perhaps indicating that M. pacificus was extirpated in the region due to habitat shifts associated with glacial encroachment. Subjects Biogeography, Evolutionary Studies, Paleontology, Zoology Keywords Mammut pacificus, Mammutidae, Montana, Irvingtonian, Pleistocene INTRODUCTION Submitted 7 May 2020 The recent recognition of the Pacific mastodon (Mammut pacificus (Dooley Jr et al., 2019)) Accepted 3 September 2020 as a new species distinct from and contemporaneous with the American mastodon Published 16 November 2020 (M. americanum) revealed an unrealized complexity in North American mammutid Corresponding author evolution during the Pleistocene. Dooley Jr et al. (2019) distinguished M. pacificus from Andrew T. McDonald, [email protected] M. americanum by a suite of dental and skeletal features: (1) upper third molars (M3) Academic editor and lower third molars (m3) much narrower relative to length in M. -

Pleistocene - History of Earth's Climate
Pleistocene - History of Earth's climate http://www.dandebat.dk/eng-klima5.htm History of Earth's Climate 5. - Cenozoic II - Pleistocene Home DH-Debate 4. Tertiary Introduction - The Pleistocene Ice Ages - The climate in the ice-free part of the World - During Last Glacial Maximum, the World became cold and 6. End of Pleistocene dusty - Temperature and CO2 - Milankovic Astronomical Climate Theory - Interglacials and other warm Periods - The Super volcano Toba - Links og literature Introduction Pleistocene is the period in Earth's history that we commonly refer to as the Ice Age. Through much of this period, the Earth's northern and southern regions were covered by kilometer thick glaciers. It is important to recognize that the Pleistocene was a series of real ice ages, separated by relatively short interglacial periods. The Pleistocene started 2.6 million years ago and lasted until the termination of the Weichsel glaciation about 11,711 years ago. Timeline of Earth's geological periods. Time progresses from right to left. The glowing inferno just after Earth was formed is named Hadean. In Archean water condensed and an atmosphere of nitrogen and methane was formed together with the first rocks that we know about. In Proterozoic cyano bacteria produced oxygen, which oxidized iron and methane, in the end of the period life emerged on the seabed. Phanerozoic represents the era in which there have been visible tangible life. It is divided in Paleozoic, Mesozoic and Cenozoic. Paleozoic was the period of early life. Mesozoic was the time of the dinosaurs, and Cenozoic is the era of mammals, which latter further is divided into Tertiary and Quaternary. -

Article Processing Charges for This Open-Access Depth, Rendering Deeper and Older OM Bioavailable for Mi- Publication Were Covered by a Research Crobial Decomposition
Biogeosciences, 15, 1969–1985, 2018 https://doi.org/10.5194/bg-15-1969-2018 © Author(s) 2018. This work is distributed under the Creative Commons Attribution 3.0 License. Substrate potential of last interglacial to Holocene permafrost organic matter for future microbial greenhouse gas production Janina G. Stapel1, Georg Schwamborn2, Lutz Schirrmeister2, Brian Horsfield1, and Kai Mangelsdorf1 1GFZ, German Research Centre for Geoscience, Helmholtz Centre Potsdam, Organic Geochemistry, Telegrafenberg, 14473 Potsdam, Germany 2Alfred Wegener Institute, Helmholtz Centre for Polar and Marine Research, Department of Periglacial Research, Telegrafenberg, A43, 14473 Potsdam, Germany Correspondence: Kai Mangelsdorf ([email protected]) Received: 14 March 2017 – Discussion started: 17 March 2017 Revised: 2 February 2018 – Accepted: 6 February 2018 – Published: 4 April 2018 Abstract. In this study the organic matter (OM) in sev- 1 Introduction eral permafrost cores from Bol’shoy Lyakhovsky Island in NE Siberia was investigated. In the context of the observed The northern areas of the Eurasian landmass are under- global warming the aim was to evaluate the potential of lain by permafrost, which is defined as ground that remains freeze-locked OM from different depositional ages to act as colder than 0 ◦C for at least 2 consecutive years (Washburn, a substrate provider for microbial production of greenhouse 1980). These areas contain a large reservoir of organic car- gases from thawing permafrost. To assess this potential, the bon freeze-locked in the permafrost deposits (French, 2007; concentrations of free and bound acetate, which form an Zimov et al., 2009). Hugelius et al. (2014) estimated that appropriate substrate for methanogenesis, were determined. -

D. Palaeozoological Research
Eiszeitalter u. Gegenwart Band 23/24 Seite 333-339 Öhringen/Württ., 15. Oktober 1973 D. Palaeozoological Research by HORST REMY, Bonn translated by U. BUREK, S. CHRULEV and R. THOMAS, Tübingen 1. Molluscs Comprehensive studies of Pleistocene land- and freshwater molluscs have so far been largely restricted to the faunas of interglacial and Würmian glacial deposits, and to the history of postglacial faunas. ANT has reconstructed the postglacial history of changes in landsnail distributions in NW-Germany and Westphalia (ANT 1963a, 1967). The present day fauna consists of a preexisting fauna ("Urfauna") together with postglacial immi grants. The elements of the "Urfauna" are eurythermal species, which were already living in NW-Germany during Würm Glaciation. These species comprise about 24 °/o of the present day fauna. In the South German periglacial region this portion is rather higher. Microclimatic conditions must have been more favourable in this region, presumably because the influence of the alpine ice mass was not as strong as that of the continental ice sheet to the north. There were many ecological niches whose local climates were more favourable for the survival of such species. Along the south coast of England, SW of the land connection with the continent, several migrant species survived (Atlanto-Britannic Fauna). In the forested region west of the Urals, the species wich today constitute the Siberio-Asiatic Fauna survived, while the immigrant species of the Mediterranean Fauna survived on the E-coast of Spain, on the W-coast of Italy and SE-Europe. It is not yet certain where some of the immigrant species lived during the glaciation; possibly these forms survived the ice age in S-Germany and the adjoining regions. -

Cypris 2016-2017
CYPRIS 2016-2017 Illustrations courtesy of David Siveter For the upper image of the Silurian pentastomid crustacean Invavita piratica on the ostracod Nymphateline gravida Siveter et al., 2007. Siveter, David J., D.E.G. Briggs, Derek J. Siveter, and M.D. Sutton. 2015. A 425-million-year- old Silurian pentastomid parasitic on ostracods. Current Biology 23: 1-6. For the lower image of the Silurian ostracod Pauline avibella Siveter et al., 2012. Siveter, David J., D.E.G. Briggs, Derek J. Siveter, M.D. Sutton, and S.C. Joomun. 2013. A Silurian myodocope with preserved soft-parts: cautioning the interpretation of the shell-based ostracod record. Proceedings of the Royal Society London B, 280 20122664. DOI:10.1098/rspb.2012.2664 (published online 12 December 2012). Watermark courtesy of Carin Shinn. Table of Contents List of Correspondents Research Activities Algeria Argentina Australia Austria Belgium Brazil China Czech Republic Estonia France Germany Iceland Israel Italy Japan Luxembourg New Zealand Romania Russia Serbia Singapore Slovakia Slovenia Spain Switzerland Thailand Tunisia United Kingdom United States Meetings Requests Special Publications Research Notes Photographs and Drawings Techniques and Methods Awards New Taxa Funding Opportunities Obituaries Horst Blumenstengel Richard Forester Franz Goerlich Roger Kaesler Eugen Kempf Louis Kornicker Henri Oertli Iraja Damiani Pinto Evgenii Schornikov Michael Schudack Ian Slipper Robin Whatley Papers and Abstracts (2015-2007) 2016 2017 In press Addresses Figure courtesy of Francesco Versino, -
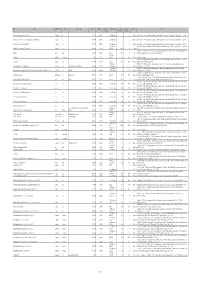
Available As PDF
ID Locality Administrative unit Country Alternate spelling Latitude Longitude Geographic Stratigraphic age Age, lower Age, upper Epoch Reference precision boundary boundary 3247 Acquasparta (along road to Massa Martana) Acquasparta Italy 42.707778 12.552333 1 late Villafranchian 2 1.1 Pleistocene Esu, D., Girotti, O. 1975. La malacofauna continentale del Plio-Pleistocene dell’Italia centrale. I. Paleontologia. Geologica Romana, 13, 203-294. 3246 Acquasparta (NE of 'km 32', below Chiesa di Santa Lucia di Burchiano) Acquasparta Italy 42.707778 12.552333 1 late Villafranchian 2 1.1 Pleistocene Esu, D., Girotti, O. 1975. La malacofauna continentale del Plio-Pleistocene dell’Italia centrale. I. Paleontologia. Geologica Romana, 13, 203-294. 3234 Acquasparta (Via Tiberina, 'km 32,700') Acquasparta Italy 42.710556 12.549556 1 late Villafranchian 2 1.1 Pleistocene Esu, D., Girotti, O. 1975. La malacofauna continentale del Plio-Pleistocene dell’Italia centrale. I. Paleontologia. Geologica Romana, 13, 203-294. upper Alluvial Ewald, R. 1920. Die fauna des kalksinters von Adelsheim. Jahresberichte und Mitteilungen des Oberrheinischen geologischen Vereines, Neue Folge, 3330 Adelsheim (Adelsheim, eastern part of the city) Adelsheim Germany 49.402305 9.401456 2 (Holocene) 0.00585 0 Holocene 9, 15-17. Sanko, A.F. 2007. Quaternary freshwater mollusks Belarus and neighboring regions of Russia, Lithuania, Poland (field guide). [in Russian]. Institute 5200 Adrov Adrov Belarus 54.465816 30.389993 2 Holocene 0.0117 0 Holocene of Geochemistry and Geophysics, National Academy of Sciences, Belarus. middle-late 4441 Adzhikui Adzhikui Turkmenistan 39.76667 54.98333 2 Pleistocene 0.781 0.0117 Pleistocene FreshGEN team decision Hagemann, J. 1976. Stratigraphy and sedimentary history of the Upper Cenozoic of the Pyrgos area (Western Peloponnesus), Greece. -

The Classical European Glacial Stages: Correlation with Deep-Sea Sediments
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by UNL | Libraries University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 1978 The Classical European Glacial Stages: Correlation with Deep-Sea Sediments George Kukla Columbia University Follow this and additional works at: https://digitalcommons.unl.edu/tnas Part of the Life Sciences Commons Kukla, George, "The Classical European Glacial Stages: Correlation with Deep-Sea Sediments" (1978). Transactions of the Nebraska Academy of Sciences and Affiliated Societies. 334. https://digitalcommons.unl.edu/tnas/334 This Article is brought to you for free and open access by the Nebraska Academy of Sciences at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Transactions of the Nebraska Academy of Sciences and Affiliated Societiesy b an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Transactions of the Nebraska Academy of Sciences- Volume VI, 1978 THE CLASSICAL EUROPEAN GLACIAL STAGES: CORRELATION WITH DEEP-SEA SEDIMENTS GEORGE KUKLA Lamont-Doherty Geological Observatory Of Columbia University Palisades, New York 10964 Four glacials and three interglacials, recognized by classical sea record revealed that this is not the case. In order to learn Alpine and North-European subdivisions of the Pleistocene, were cor the reasons for the discrepancy, we identified the type units related with continuous oxygen-isotope records from the oceans using and their boundaries at the type localities, correlated them loess sections and terraces as a link (Fig. 15). It was found that the Alpine "glacial" stages are represented by sediments formed during with the standard stratigraphic system, and tried to recognize both glacial and interglacial climates, that the classical Alpine "inter their true climatic characteristics. -

Downloaded From
A continent-wide framework for local and regional stratigraphies Gijssel, K. van Citation Gijssel, K. van. (2006, November 22). A continent-wide framework for local and regional stratigraphies. Retrieved from https://hdl.handle.net/1887/4985 Version: Not Applicable (or Unknown) Licence agreement concerning inclusion of doctoral thesis in the License: Institutional Repository of the University of Leiden Downloaded from: https://hdl.handle.net/1887/4985 Note: To cite this publication please use the final published version (if applicable). Chapter 4 A SUPPLEMENTARY STRATIGRAPHICAL FRAMEWORK FOR NORTHWEST AND CENTRAL EU- ROPE ON THE BASIS OF GENETIC SEQUENCE AND EVENT STRATIGRAPHY Having reviewed the contemporary Middle Pleistocene stratigra- Basin which have acted as main Pleistocene depocentres, such phy of Northwest and Central Europe by discussing five broad as the Central Graben, the Sole Pit and three composed subba- categories of environments and their sedimentary products, they sins in the southern part of the North Sea Basin: the Anglo- are now placed into a framework of interregional extent and sig- Dutch (Broad Fourteens, Western Netherlands), the North Ger- nificance. Such a large-scale framework requires a material basis man and the Polish sub-basins. The eastern part of the North- from the type localities and type regions with uniformly defined west European Basin was only marginally influenced by tecton- units for interpretation. Since the existing (national) classification ics during the Pleistocene. systems are based on different criteria, a supplementary, non-in- - The continued activity of rift structures in the Central European terpretive stratigraphical framework is advocated in this chapter in uplands in between the North Sea Basin and the Alps (Ziegler which the existing litho-, bio-, soil- and other stratigraphical ele- 1994). -

The Stratigraphic Table of Germany 2016 (STG 2016)
Menning, M. (2018): Die Stratigraphische Tabelle von Deutschland 2016 (STD 2016). The Stratigraphic Table of Germany 2016 (STG 2016). - Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 169, 2, 105-128. https://doi.org/10.1127/zdgg/2018/0161 Institional Repository GFZpublic: https://gfzpublic.gfz-potsdam.de/ 1 Die Stratigraphische Tabelle von Deutschland 2016 (STD 2016) / The Stratigraphic Table of Germany 2016 (STG 2016) Manfred Menning *Anschrift der Autors: Dr. Manfred Menning, Helmholtz-Zentrum Potsdam, Deutsches GeoForschungsZentrum GFZ, Telegraphenberg, 14473 Potsdam ([email protected]) Kurzfassung Intention und Realität der Stratigraphischen Tabelle von Deutschland 2016 (STD 2016) werden dargestellt. Den Zeitabschnitten Ediacarium bis Silur sowie Späte Trias bis Quartär liegt die Zeitskala der GTS 2012 (Geological Time Scale 2012) zu Grunde. Für das Devon wurde eine eigene Zeitskala entwickelt, für Karbon und Perm wird die Zeitskala der Stratigraphischen Tabelle von Deutschland 2002 (STD 2002) genutzt und für die Frühe und Mittlere Trias die Zeitskala der Erläuterungen 2005 zur Stratigraphischen Tabelle von Deutschland 2002 (ESTD 2005). Die Alter der paläozoischen und mesozoischen Stufen sind auf 0,5 Ma bzw. 1 Ma gerundet. Die zeitliche Einstufung vieler mitteleuropäischer Einheiten in die Globale Stratigraphische Skala ist variabel und nicht endgültig. Dies verdeutlichen Pfeile an zahlreichen Schichtgrenzen. Dargestellt sind möglichst viele gebräuchliche Schichten mit ihren aktuellen Namen und zusätzlich auch oft mit ihren eingebürgerten, aber inzwischen obsoleten Namen. Umstritten ist die Existenz nahezu beckenweiter Schichtlücken, so im Keuper, während solche Lücken im höheren Rotliegend und an der Kreide-Tertiär-Grenze auch im Norddeutschen Becken weithin akzeptiert werden. Neuigkeiten gegenüber der STD 2002 und ausgewählte, darunter grundlegende stratigraphische Probleme sowie Besonderheiten in den einzelnen Zeitabschnitten der STD 2016 werden diskutiert. -

The Occurrence 1834) in Britain. of Micrasema Setiferum
© Hans Malicky/Austria; download unter www.biologiezentrum.at 21 BRAUERIA (Lunz am See, Austria) 35:21-22 (2008) VERNEAUX (1972) describes the ecology of this species in the Jura Mountains where the larvae, often occurring in large The occurrence of Micrasema setiferum (PlCTET numbers, colonise mosses in flows of+/- 20-70 cm/sec and at 1834) in Britain. water depths of 25-60 cm. In the River Doubs, where much of this study was undertaken, the water temperature ranged from 1.4-21.6 °C, an annual range of 20.2 °C. M.T.GREENWOOD The present distribution of Micrasema setiferum is shown in This note is to supplement the data presented by GREEN et al GREENWOOD et al (2003). In this paper, the present (2006), following an extensive multidisciplinary study of, geographical distribution has been converted into climate what are considered to be fluvial deposits from a short time space, this being described by plotting the mean temperature interval within a late Middle Pleistocene interglacial (Marine of the warmest month (Tmax) against the difference between Isotope Stage 9). From the organic sands and silts at the mean temperature of the coldest month and that of the Hackney, north London, the plant, mollusc and insect warmest month, usually July and January, in °C (Trange). (Colcoptera) assemblages, all suggest a climate more Using this procedure, the conditions tolerated by M. setiferum continental than that found in southern Britain today. fall within a climate envelop of greater continentality, than exists in the Trent valley today. In the course of preparing material for the above paper, Professor G.R.Coope, having concentrated on fossil The significance of this find in MIS9 is that it is the oldest Coleoptera, also collected frontoclypeal, pronotal and record of this species in Britain and of only the third known mesonotal fragments of larval Trichoptera, sending these to locality.