An Experiment-Based Methodology for Classical Genetics and Molecular Biology
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Introduction to Genetics Bios 225
INTRODUCTION TO GENETICS BIOS 225 Course Description This course introduces the student to the basic concepts of inheritance, populations, mutations, and techniques used to assess each of these. Credit: 2 credits Repeatable: No Course Structure The course will be presented in different formats: Lectures with PowerPoints, self-directed learning, discussions and student assignments etc. Competencies This course emphasizes competencies to enhance skills essential for a future health care professional. • Knowledge o Demonstrate content knowledge and skills in foundational courses required by biomedical professionals o Demonstrate information literacy o Demonstrate quantitative reasoning o Demonstrate longitudinal learning through coursework • Critical Thinking o Develop the skills of self-reflection and peer assessment to improve personal performance. o Demonstrate the ability to analyze literature and written material o Demonstrate the ability to distinguish between well-reasoned and poorly reasoned arguments • Communication Skills o Demonstrate effective presentation skills to faculty and peers. o Demonstrate effective listening skills o Demonstrate effective written communication 1 Objectives: Upon completion of BIOS 225 course, the student should be able to describe: • The structure and function of purines, pyrimidines, nucleosides and nucleotides • The structure and functions of nucleic acids (DNA and RNA) • The chromosome anatomy and human karyotypes • The concepts of prokaryotic and eukaryotic DNA replication • The concepts of prokaryotic and eukaryotic RNA transcription and post-transcriptional modifications • The concepts of prokaryotic and eukaryotic protein translation and post-translational modifications • The regulations of prokaryotic and eukaryotic gene expressions • The process of genomic, chromosomal and gene mutations; and its repair mechanism • The Mendel’ hypothesis and molecular mechanisms of genetic inheritance Schedule: Dates and times to be posted at the beginning of the term on the online calendar. -

Models of the Gene Must Inform Data-Mining Strategies in Genomics
entropy Review Models of the Gene Must Inform Data-Mining Strategies in Genomics Łukasz Huminiecki Department of Molecular Biology, Institute of Genetics and Animal Biotechnology, Polish Academy of Sciences, 00-901 Warsaw, Poland; [email protected] Received: 13 July 2020; Accepted: 22 August 2020; Published: 27 August 2020 Abstract: The gene is a fundamental concept of genetics, which emerged with the Mendelian paradigm of heredity at the beginning of the 20th century. However, the concept has since diversified. Somewhat different narratives and models of the gene developed in several sub-disciplines of genetics, that is in classical genetics, population genetics, molecular genetics, genomics, and, recently, also, in systems genetics. Here, I ask how the diversity of the concept impacts data-integration and data-mining strategies for bioinformatics, genomics, statistical genetics, and data science. I also consider theoretical background of the concept of the gene in the ideas of empiricism and experimentalism, as well as reductionist and anti-reductionist narratives on the concept. Finally, a few strategies of analysis from published examples of data-mining projects are discussed. Moreover, the examples are re-interpreted in the light of the theoretical material. I argue that the choice of an optimal level of abstraction for the gene is vital for a successful genome analysis. Keywords: gene concept; scientific method; experimentalism; reductionism; anti-reductionism; data-mining 1. Introduction The gene is one of the most fundamental concepts in genetics (It is as important to biology, as the atom is to physics or the molecule to chemistry.). The concept was born with the Mendelian paradigm of heredity, and fundamentally influenced genetics over 150 years [1]. -

Epigenetics Is Normal Science, but Don't Call It Lamarckian
Extended Abstract Volume 6:3, 2020 DOI: 6.2472/imedipce.2020.6.002 Journal of Clinical Epigenetics ISSN: 2472-1158 Open Access Epigenetics is Normal Science, But Don’t Call It Lamarckian David Penny New Zealand Extended Abstract they are general (but not necessarily universal) because they are found in Excavates, which many people consider are ancestral to other eukaryotes [6, 7]. Similarly, an important point is that Epigenetic mechanisms are increasingly being accepted as an integral methylation, phosphorylation, etc. of the DNA and of the histones is a part of normal science. Earlier genetic studies appear to have part of normal evolution (at least in eukaryotes that have the assumed that the only changes were ‘mutations’ as a result of a nucleosome structure) [8]. One of the most interesting aspects is change in the DNA sequence. If any such changes altered the amino methylation (and hydroxymethylation) of cytosine, and this is found in acid sequences then it was effectively a mutation, and if it occurred in many deep branching eukaryotes [9]. Typically there is methylation of a germline cell (including in unicellular organisms) then it could be cytosine molecules, especially in CpG (cytosine followed by guanine) passed on to subsequent generations. However, it is increasingly regions. The role of small RNAs in helping regulate the levels of apparent that other changes are also important, for example, changes protein expression is discussed in [10]. The main point here is that to methylation patterns, and to the histone proteins. Epigenetics was protein expression levels are very variable in all cells – it is not just sometimes consider ‘additional’ to classical genetics, but it is still fully classical genetics that is important; there are many epigenetic dependent on DNA, RNA and protein sequences, and at least since [1] mechanisms affecting the differences in protein expression. -

Using Classical Genetics Simulator (CGS) to Teach Students the Basics of Genetic Research
Tested Studies for Laboratory Teaching Proceedings of the Association for Biology Laboratory Education Vol. 33, 85–94, 2012 Using Classical Genetics Simulator (CGS) to Teach Students the Basics of Genetic Research Jean Heitz1, Mark Wolansky2, and Ben Adamczyk3 1University of Wisconsin, Zoology Department, 250 N. Mills Street, Madison WI 53706 USA 2University of Alberta, Department of Biological Science, Edmonton AB, CAN T6G 2E9 3Genedata, MA, A-1 Cranberry HI, Lexington MA 02421 USA ([email protected];[email protected];[email protected]) Some experiments are not well suited to large introductory labs because of space, time and funding constraints. Among these are classical Mendelian genetics investigations. Cyber labs can overcome these constraints. Classi- cal Genetics Simulator (CGS) gives students the opportunity to perform controlled crosses with model organisms much like a geneticist would do. CGS provides populations of Drosophila, Arabidopsis or mice with unknown patterns of inheritance and gives students the tools to design and perform experiments to discover these patterns. Students require an understanding of the what, why and how of solving genetic problems to successfully complete our cyber genetics lab activities. Keywords: Mendelian genetics, simulator, classical genetics simulator (CGS), linkage, monohybrid, dihybrid Introduction In large introductory biology classes some labs are dif- How can students get practice doing what geneticists ficult to do because of space, time and funding constraints. actually do? Among these are classical Mendelian genetics investiga- An obvious way for students to get practice doing what tions. As a result, we have turned to cyber labs. Working geneticists do is to conduct actual crosses. -

Animal Traditions: Behavioural Inheritance in Evolution
Animal Traditions: Behavioural Inheritance in Evolution Eytan Avital and Eva Jablonka CAMBRIDGE UNIVERSITY PRESS Animal Traditions Behavioural Inheritance in Evolution Animal Traditions maintains that the assumption that the selection of genes supplies both a sufficient explanation of the evolution of behav- iour and a true description of its course is, despite its almost univer- sal acclaim, wrong. Eytan Avital and Eva Jablonka contend that evolutionary explanations must take into account the well-established fact that, in mammals and birds, the transfer of learnt information across generations is both ubiquitous and indispensable. The introduc- tion of the behavioural inheritance system into the Darwinian explanatory scheme enables the authors to offer new interpretations for common behaviours such as maternal behaviours, behavioural conflicts within families, adoption and helping. This approach offers a richer view of heredity and evolution, integrates developmental and evolutionary processes, suggests new lines for research and provides a constructive alternative to both the selfish gene and meme views of the world. It will make stimulating reading for all those interested in evolutionary biology, sociobiology, behavioural ecology and psychology. eytan avital is a lecturer in Zoology in the Department of Natural Sciences at David Yellin College of Education in Jerusalem. He is a highly experienced field biologist, and has written one zoology text and edited several others on zoology and evolution for the Israel Open university. eva jablonka is a senior lecturer in the Cohn Institute for the History and Philosophy of Science and Ideas, at Tel-Aiv University. She is the author of three books on heredity and evolution, most recently Epigenetic Inheritance and Evolution with Marion Lamb. -

The Sustained Impact of Model Organisms—In Genetics and Epigenetics
| COMMENTARY The Sustained Impact of Model Organisms—in Genetics and Epigenetics Nancy M. Bonini*,†,1 and Shelley L. Berger‡,1 *Department of Cellular and Developmental Biology, Perlman School of Medicine, †Department of Biology, and ‡Department of Genetics, University of Pennsylvania, Philadelphia, Pennsylvania 19104 KEYWORDS classical genetics; epigenetics; behavior; genomics; human disease Now that DNA sequencing can reveal disease-relevant muta- The Rise of Model Organism Genetics and the tions in humans, it is appropriate to consider the history of Embrace of Molecular Genetics genetic research on model organisms, and whether model Model organism genetics has revealed many fundamental organisms will continue to play an important role. Our view is biological processes. This was achieved by applying unbiased that genetic research in model organisms has had an in- genetic screens to reveal genes and their mechanisms of disputably enormous impact on our understanding of com- action in processes such as cell cycle control (Nasmyth plex biological pathways and their epigenetic regulation, and 2001), basic body plan specification and mechanisms of de- on the development of tools and approaches that have been velopment (Lewis 1978; Nusslein-Volhard and Wieschaus crucial for studies of human biology. We submit that model 1980), and cell death pathways (Ellis and Horvitz 1986; organisms will remain indispensable for interpreting complex Avery and Horvitz 1987), all of which were recognized with genomic data, for testing hypotheses regarding function, and Nobel prizes. for further study of complex behaviors, to reveal not only The sequencing of genomes revealed the breadth of the novel genetic players, but also the profound impact of epige- conservation of genes, and of entire pathways. -
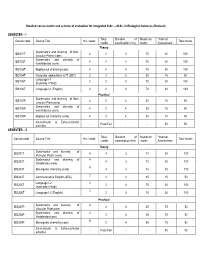
Detailed Course Matrix and Scheme of Evaluation for Integrated B.Sc. – M.Sc. in Biological Sciences (Revised) SEMESTER –
Detailed course matrix and scheme of evaluation for Integrated B.Sc. – M.Sc. in Biological Sciences (Revised) SEMESTER – I Total Duration of Maximum *Internal Course code Course Title Hrs / week Total marks credits examination (hrs) marks Assessment Theory Systematics and diversity of Non- IBS101T 4 4 3 70 30 100 vascular Plants (core) Systematics and diversity of IBS102T 4 4 3 70 30 100 Invertebrates (core) IBS103T Biophysical chemistry(core) 4 4 3 70 30 100 IBS104T Computer applications & IT (SEC) 2 2 2 35 15 50 Language I-1 IBS105T 2 2 3 70 30 100 (Kannada / Hindi) IBS106T Language I-1 (English) 2 2 3 70 30 100 Practical Systematics and diversity of Non- IBS107P 4 2 4 35 15 50 vascular Plant (core) Systematics and diversity of IBS108P 4 2 4 35 15 50 Invertebrates (core) IBS109P Biophysical chemistry (core) 4 2 4 35 15 50 Co-curricular & Extra-curricular - Pass/Fail - - 50 50 activities SEMESTER – II Total Duration of Maximum *Internal Course code Course Title Hrs / week Total marks credits examination (hrs) marks Assessment Theory Systematics and diversity of IBS201T 4 4 3 70 30 100 Vascular Plants (core) Systematics and diversity of 4 IBS202T 4 3 70 30 100 Vertebrates (core) 4 IBS203T Bio-organic chemistry (core) 4 3 70 30 100 2 IBS204T Communicative English (AEC) 2 2 35 15 50 Language I-2 2 IBS205T 2 3 70 30 100 (Kannada / Hindi) 2 IBS206T Language I-2 (English) 2 3 70 30 100 Practical Systematics and diversity of 4 IBS207P 2 4 35 15 50 Vascular Plant(core) Systematics and diversity of 4 IBS208P 2 4 35 15 50 Vertebrates(core) 4 IBS209P -

INTRODUCTION to GENETICS Table of Contents Heredity, Historical
INTRODUCTION TO GENETICS Table of Contents Heredity, historical perspectives | The Monk and his peas | Principle of segregation Dihybrid Crosses | Mutations | Genetic Terms | Links Heredity, Historical Perspective | Back to Top For much of human history people were unaware of the scientific details of how babies were conceived and how heredity worked. Clearly they were conceived, and clearly there was some hereditary connection between parents and children, but the mechanisms were not readily apparent. The Greek philosophers had a variety of ideas: Theophrastus proposed that male flowers caused female flowers to ripen; Hippocrates speculated that "seeds" were produced by various body parts and transmitted to offspring at the time of conception, and Aristotle thought that male and female semen mixed at conception. Aeschylus, in 458 BC, proposed the male as the parent, with the female as a "nurse for the young life sown within her". During the 1700s, Dutch microscopist Anton van Leeuwenhoek (1632-1723) discovered "animalcules" in the sperm of humans and other animals. Some scientists speculated they saw a "little man" (homunculus) inside each sperm. These scientists formed a school of thought known as the "spermists". They contended the only contributions of the female to the next generation were the womb in which the homunculus grew, and prenatal influences of the womb. An opposing school of thought, the ovists, believed that the future human was in the egg, and that sperm merely stimulated the growth of the egg. Ovists thought women carried eggs containing boy and girl children, and that the gender of the offspring was determined well before conception. -

2014-2015 and Is Accurate and Current, to the Greatest Extent Possible, As of June 2014
Cover Cover 1 University’s Mission Statement James B. Duke’s founding Indenture of Duke University directed the members of the University to “provide real leadership in the educational world” by choosing individuals of “outstanding character, ability and vision” to serve as its officers, trustees and faculty; by carefully selecting students of “character, determination and application;” and by pursuing those areas of teaching and scholarship that would “most help to develop our resources, increase our wisdom and promote human happiness.” To these ends, the mission of Duke University is to provide a superior liberal education to undergraduate students, attending not only to their intellectual growth but also to their development as adults committed to high ethical standards and full participation as leaders in their communities; to prepare future members of the learned professions for lives of skilled and ethical service by providing excellent graduate and professional education; to advance the frontiers of knowledge and contribute boldly to the international community of scholarship; to promote an intellectual environment built on a commitment to free and open inquiry; to help those who suffer, cure disease and promote health, through sophisticated medical research and thoughtful patient care; to provide wide ranging educational opportunities, on and beyond our campuses, for traditional students, active professionals and life-long learners using the power of information technologies; and to promote a deep appreciation for the range of human difference and potential, a sense of the obligations and rewards of citizenship, and a commitment to learning, freedom and truth. By pursuing these objectives with vision and integrity, Duke University seeks to engage the mind, elevate the spirit, and stimulate the best effort of all who are associated with the University; to contribute in diverse ways to the local community, the state, the nation and the world; and to attain and maintain a place of real leadership in all that we do. -

G14TBS Part II: Population Genetics
G14TBS Part II: Population Genetics Dr Richard Wilkinson Room C26, Mathematical Sciences Building Please email corrections and suggestions to [email protected] Spring 2015 2 Preliminaries These notes form part of the lecture notes for the mod- ule G14TBS. This section is on Population Genetics, and contains 14 lectures worth of material. During this section will be doing some problem-based learning, where the em- phasis is on you to work with your classmates to generate your own notes. I will be on hand at all times to answer questions and guide the discussions. Reading List: There are several good introductory books on population genetics, although I found their level of math- ematical sophistication either too low or too high for the purposes of this module. The following are the sources I used to put together these lecture notes: • Gillespie, J. H., Population genetics: a concise guide. John Hopkins University Press, Baltimore and Lon- don, 2004. • Hartl, D. L., A primer of population genetics, 2nd edition. Sinauer Associates, Inc., Publishers, 1988. • Ewens, W. J., Mathematical population genetics: (I) Theoretical Introduction. Springer, 2000. • Holsinger, K. E., Lecture notes in population genet- ics. University of Connecticut, 2001-2010. Available online at http://darwin.eeb.uconn.edu/eeb348/lecturenotes/notes.html. • Tavar´eS., Ancestral inference in population genetics. In: Lectures on Probability Theory and Statistics. Ecole d'Et´esde Probabilit´ede Saint-Flour XXXI { 2001. (Ed. Picard J.) Lecture Notes in Mathematics, Springer Verlag, 1837, 1-188, 2004. Available online at http://www.cmb.usc.edu/people/stavare/STpapers-pdf/T04.pdf Gillespie, Hartl and Holsinger are written for non-mathematicians and are easiest to follow. -

Mechanisms and Consequences of Epigenetic Inheritance Following Parental Preconception Alcohol Exposure
Loyola University Chicago Loyola eCommons Dissertations Theses and Dissertations 2018 Mechanisms and Consequences of Epigenetic Inheritance Following Parental Preconception Alcohol Exposure Annadorothea Asimes Loyola University Chicago Follow this and additional works at: https://ecommons.luc.edu/luc_diss Part of the Neurosciences Commons Recommended Citation Asimes, Annadorothea, "Mechanisms and Consequences of Epigenetic Inheritance Following Parental Preconception Alcohol Exposure" (2018). Dissertations. 2770. https://ecommons.luc.edu/luc_diss/2770 This Dissertation is brought to you for free and open access by the Theses and Dissertations at Loyola eCommons. It has been accepted for inclusion in Dissertations by an authorized administrator of Loyola eCommons. For more information, please contact [email protected]. Copyright © 2018 Annadorothea Asimes LOYOLA UNIVERSITY CHICAGO MECHANISMS AND CONSEQUENCES OF EPIGENETIC INHERITANCE FOLLOWING PARENTAL PRECONCEPTION ALCOHOL EXPOSURE A DISSERTATION SUBMITTED TO THE FACULTY OF THE GRADUATE SCHOOL IN CANDIDACY FOR THE DEGREE OF DOCTOR OF PHILOSOPHY PROGRAM IN NEUROSCIENCE BY ANNADOROTHEA ASIMES CHICAGO, IL MAY 2018 Copyright by AnnaDorothea Asimes, 2018 ALL rights reserved. ACKNOWLEDGEMENTS The work required to compLete this dissertation wouLd not have been possibLe alone. I wouLd first Like to thank my advisor and mentor, Dr. Toni Pak. The guidance and support you have given me throughout these graduate schooL years has been invaluabLe. You have provided a chalLenging environment that pushed me to be resourcefuL, creative, boLd, and humbLed. I wouLd not be the scientist I am today without your mentorship and friendship. I wouLd also Like to thank my Lab mates, who quickLy turned from coworkers to friends. I Look forward to buiLding our Lab famiLy and seeing the great work you alL go on to do. -

Mendelian Gene ” and the ” Molecular Gene ” : Two Relevant Concepts of Genetic Units V Orgogozo, Alexandre E
The ” Mendelian Gene ” and the ” Molecular Gene ” : Two Relevant Concepts of Genetic Units V Orgogozo, Alexandre E. Peluffo, Baptiste Morizot To cite this version: V Orgogozo, Alexandre E. Peluffo, Baptiste Morizot. The ” Mendelian Gene ” and the ”Molec- ular Gene ” : Two Relevant Concepts of Genetic Units. Virginie Orgogozo. Genes and Evolu- tion, 119, Elsevier, pp.1-26, 2016, Current Topics in Developmental Biology, 978-0-12-417194-7. 10.1016/bs.ctdb.2016.03.002. hal-01354346 HAL Id: hal-01354346 https://hal.archives-ouvertes.fr/hal-01354346 Submitted on 19 Aug 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. ARTICLE IN PRESS The “Mendelian Gene” and the “Molecular Gene”: Two Relevant Concepts of Genetic Units V. Orgogozo*,1, A.E. Peluffo*, B. Morizot† *Institut Jacques Monod, UMR 7592, CNRS-Universite´ Paris Diderot, Sorbonne Paris Cite´, Paris, France †Universite´ Aix-Marseille, CNRS UMR 7304, Aix-en-Provence, France 1Corresponding author: e-mail address: [email protected] Contents 1. Introduction 2 2. The “Mendelian Gene” and the “Molecular Gene” 5 3. Current Literature Often Confuses the “Mendelian Gene” and the “Molecular Gene” Concepts 7 4.