A Faunal Analysis of Okrn-1, Banks Island, N.W.T
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Tipi Instructions
Tipi Instructions Hello and thanks for buying one of our Tipi Tents. Here are the instructions for the best way to put it up, take it down and how to store when not in use. We’ve provided a step-by-step picture guide for you to follow along with the written instructions. At the bottom of these instructions we’ve added some advice and tips on how to get years of good service out of your tent by looking after it properly. Enjoy! And don’t hesitate to contact us if you have any questions, good ideas or better ways of doing it. Once you’ve erected your tent, we would love to see your Boutique Camping pictures, so please send them to us at [email protected]. They may even feature on our blog or Facebook page! Putting up a Tipi Tent Some general advice • We advise having 2 people to erect the tent, as it will of course simplify the process. By all means, you can put it up on your own, but of course the more the merrier, easier and quicker! • Find a flat piece of land, with ample space to comfortably put up your tent remembering to include room for the guy ropes • To prevent damaging the groundsheet, remove all sharp objects from the area. Things like stones and roots etc • Be careful when creating tension with the guy ropes, always try to keep the tension even, and never over do it. Be firm but gentle. • Before you peg out the tent, make sure all zips are closed, and check again when you adjust the guy ropes • Peg out the guy ropes in line with the tent seams How to Put Up Your Tipi A1)Open all bags and lay out all parts. -
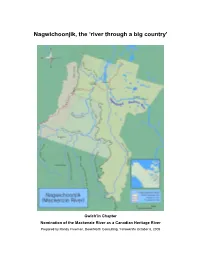
Gwich'in Nomination Chapter
Nagwichoonjik, the ‘river through a big country’ Gwich’in Chapter Nomination of the Mackenzie River as a Canadian Heritage River Prepared by Randy Freeman, DownNorth Consulting, Yellowknife October 6, 2005 2 The Mackenzie River corridor from Vihtr’ii tshik [Thunder River] to about Srehtadhadlaii [Point Separation] … was important for a number of reasons. It was one of the links connecting the different regions, and most families travelled on the river at some point during the year. Many families also spent the summer season at one of the fish camps located on both sides of the river. The sites for the great summer gatherings of ts’ii deii days were also located along this stretch of the river. So important is the river that, in [Gwich’in Elder] Gabe Andre’s words, it is a ‘highway’ of the Gwichya Gwich’in.1 1 Michael Heine, Alestine Andre, Ingrid Kritsch, Alma Cardinal and the Elders of Tsiigehtchic. Gwichya Gwich’in Googwandak: The History and Stories of the Gwichya Gwich’in. (Tsiigehtchic and Yellowknife, NT: Gwich’in Social and Cultural Institute, 2001) 59. Nagwichoonjik, the ‘river through a big country’ 3 Introduction Nagwichoonjik is the traditional Gwich’in name for the Mackenzie River. The name means ‘river through a big country’ and for the Gwich’in of the Northwest Territories it’s both literally and figuratively their main ‘highway’, the backbone of their traditional lands. Background The Gwich’in inhabit an expanse of sub-arctic boreal forest stretching from the interior of Alaska eastward through northern Yukon to the lower Mackenzie valley of the Northwest Territories. -

The Sámi People and Their Culture the Sámi Or Saami Were Also Called Lapps Or Laplanders by the English
The Sámi people and their culture The Sámi or Saami were also called Lapps or Laplanders by the English. Sámi people consider the English terms derogatory. The Sámi are recognized as the only indigenous people of Europe. They have lived in Norway, Sweden, Finland and Russia. Their origins are Finno‐Ugric, a Hungarian and Yugra (Urals) past, inhabiting the Sápmi region. Today, the region encompasses large parts of Norway and Sweden, northern parts of Finland, and the Murmansk Oblast (Kola Peninsula) of Russia. The Sámi people have their own language, culture and customs that differ from others around them. This has caused the Sámi social problems and culture clashes. As we learned from our Sámi culture presentation and a quote from ‐religiousstudiesproject.com the following: “The history between the Sámi and the Norwegian government has left a stain on the Sámi for generations: The Norwegianization policy undertaken by the Norwegian government from the 1850s up until the Second World War resulted in the apparent loss of Sami language and assimilation of the coastal Sami as an ethnically‐distinct people into the northern Norwegian population. Together with the rise of an ethno‐political movement since the 1970s, however, Sami culture has seen a revitalization of language, cultural activities, and ethnic identity (Brattland 2010:31).” Note: Suggested readings, ‐laits.utexas.edu, a 19‐part series by the University of Texas entitled “Sámi Culture.” The other reading is‐ unsr.vtaulicorpuz.org. It is a report by the United Nations on the rights of indigenous people such as the Sámi. Reindeer are the Sámi key element to how they live. -

INUVIALUIT LANGUAGE and IDENTITY: PERSPECTIVES on the SYMBOLIC MEANING of INUVIALUKTUN in the CANADIAN WESTERN ARCTIC by Alexand
INUVIALUIT LANGUAGE AND IDENTITY: PERSPECTIVES ON THE SYMBOLIC MEANING OF INUVIALUKTUN IN THE CANADIAN WESTERN ARCTIC by Alexander C. Oehler B.A., University of Northern British Columbia, 2010 THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF ARTS IN INTERDISCIPLINARY STUDIES UNIVERSITY OF NORTHERN BRITISH COLUMBIA July 2012 © Alexander C. Oehler, 2012 Abstract: The revitalization of ancestral languages has been an issue of great concern to Aboriginal communities across North America for several decades. More recently, this concern has also found a voice in educational policy, particularly in regions where Aboriginal land claims have been ratified, and where public schools fall under a mandate to offer curricula that meet the needs of Aboriginal students. This research seeks to explore the cultural significance of Inuvialuktun, a regional Inuit language comprised of three distinct dialects traditionally spoken by the Inuvialuit of the northern Northwest Territories, Canada. More specifically, the research seeks to examine the role of current Inuvialuktun language revitalization efforts in the establishment of Inuvialuit collective and individual identities across several age groups. Tying into the sociolinguistic discourse on ancestral language revitalization in North America, the research seeks to contribute a case study from a region underrepresented in the literature on language and identity. The applied aim of the study is to provide better insight on existing language ideologies and language attitudes subscribed to by current and potential learners of Inuvialuktun in the community of Inuvik, NWT. Data obtained by the study is intended to aid local and territorial language planners in identifying potential obstacles and opportunities regarding language learner motivation. -

Inuit Knowledge and Use of Wood Resources on the West Coast Of
Document generated on 10/02/2021 5:05 a.m. Études/Inuit/Studies Inuit knowledge and use of wood resources on the west coast of Nunavik, Canada Savoir inuit et utilisation des ressources en bois sur la côte ouest du Nunavik, Canada Stéphanie Steelandt, Najat Bhiry, Dominique Marguerie, Caroline Desbiens, Minnie Napartuk and Pierre M. Desrosiers Volume 37, Number 1, 2013 Article abstract Driftwood and shrubs are the primary wood resources available in most areas URI: https://id.erudit.org/iderudit/1025259ar of coastal Nunavik. Today, they are mainly used as fuel for campfires, but DOI: https://doi.org/10.7202/1025259ar historically they were very important for the ancestors of present-day Inuit. This article documents Inuit traditional knowledge about the origin, See table of contents availability, gathering, and exploitation of wood resources in this region located in the Low Arctic and the Subarctic. Interviews were conducted with 27 Inuit between 60 and 89 years of age in the villages of Ivujivik, Akulivik, Publisher(s) Inukjuak, and Umiujaq on the east coast of Hudson Bay. Our data reveal, among other things, that Inuktitut names for pieces of driftwood were based Association Inuksiutiit Katimajiit Inc. on shape, aspect, colour, and texture. This traditional knowledge was very Centre interuniversitaire d’études et de recherches autochtones (CIÉRA) accurate and highly diverse in the southern villages because of their significant exposure to driftwood. Wood from shrubs (i.e. willows, birches, and alders) ISSN was mainly harvested in the fall and used to make fires, mattresses, sleeping mats (alliat), and other objects. According to the participants, driftwood 0701-1008 (print) originates in southern Hudson Bay and James Bay and is washed up on the 1708-5268 (digital) beaches in late summer and the fall. -

Tent Inspection
Tent Guide Information referenced from the 2015 Frederick County Fire Prevention Code A Tent is defined as a structure, enclosure or shelter, with or without sidewalls or drops, constructed of fabric or pliable material supported by any manner except by air or the contents that it protects. All Tents: • A permit issued by the Frederick County Fire Marshal’s office shall be required for tents over 900 square feet not being used for recreational camping purposes. (Frederick County fire Prevention Code section 107.2 Permit Required.) • A detailed site and floor plan for tents with occupant load of 50 or more shall be required with each application for approval. The tent floor plan shall indicate details of means of egress, seating capacity, arrangement of seating and location and types of heating and electrical equipment. (Frederick County fire Prevention Code section 3103.6 Construction documents.) • An unobstructed fire break passageway or fire road not less than 12 feet wide and free from guy ropes or other obstructions shall be maintained on all sides of all tents unless otherwise approved by the fire code official. (Frederick County fire Prevention Code section 3103.8.6 Fire break.) • A certificate from an approved laboratory stating that the materials used in tent meet the flame- retardant criteria needed to pass NFPA Test Method 1 or Test Method 2. (Frederick County fire Prevention Code section 3104.2 Flame propagation performance treatment.) • Approved “No Smoking” signs shall be conspicuously posted. (Frederick County fire Prevention Code section 3104.6 Smoking.) • At least one 5lb. multipurpose 2A 10BC portable fire extinguisher shall be hung and tagged within 75 ft of travel distance. -

The Four-Year Arctic Odyssey of HMS Investigator and the Mcclure
REVIEWS • 109 DISCOVERING THE NORTH-WEST PASSAGE: THE FOUR-YEAR ARCTIC ODYSSEY OF H.M.S. INVESTIGATOR AND THE McCLURE EXPEDITION. By GLENN M. STEIN. Jefferson, North Carolina: McFarland, 2015. ISBN 978-0-7864-7708-1. maps, b&w illus., appendices, notes, bib., index. Available from www.mcfarlandpub.bom; order line: 800-253-2187. Softbound. US$39.95. Of all the explorers who sailed from Europe in search of the Northwest Passage, the least admirable was probably Robert McClure. He is remembered today because in 1855, the British government awarded him a knighthood and 10 000 pounds (today worth roughly $1.3 million), with half going to his men for having become “the first to have passed by water from sea to sea.” By returning alive to Eng- land, they had provided “a living evidence of the existence of a North-west Passage” (p. 240). Originally the reward had been offered, as author Glenn M. Stein notes, “for the first person who should complete the North-west Passage, by actually sailing with his ship from one ocean to the other” (p. 240). But the government, saddled with the expenses of the Crimean War, was desper- ate to staunch the spending on Arctic search expeditions. And so it recognized McClure’s tortured logic, turning a blind eye to the fact that, as Stein writes, “the Investigators had walked through portions of the route” (p. 240). In Discovering the North-West Passage, author Stein details the difficult human story behind McClure’s dubi- ous achievement. This is the definitive account of the last voyage of HMS Investigator—a tale of how a Machiavel- lian captain, obsessive and ruthless, turned his ship into a frozen hell of unjust floggings, imprisonments, starvation, scurvy, and even, for a few, miserable deaths. -

Frederick J. Krabbé, Last Man to See HMS Investigator Afloat, May 1854
The Journal of the Hakluyt Society January 2017 Frederick J. Krabbé, last man to see HMS Investigator afloat, May 1854 William Barr1 and Glenn M. Stein2 Abstract Having ‘served his apprenticeship’ as Second Master on board HMS Assistance during Captain Horatio Austin’s expedition in search of the missing Franklin expedition in 1850–51, whereby he had made two quite impressive sledge trips, in the spring of 1852 Frederick John Krabbé was selected by Captain Leopold McClintock to serve under him as Master (navigation officer) on board the steam tender HMS Intrepid, part of Captain Sir Edward Belcher’s squadron, again searching for the Franklin expedition. After two winterings, the second off Cape Cockburn, southwest Bathurst Island, Krabbé was chosen by Captain Henry Kellett to lead a sledging party west to Mercy Bay, Banks Island, to check on the condition of HMS Investigator, abandoned by Commander Robert M’Clure, his officers and men, in the previous spring. Krabbé executed these orders and was thus the last person to see Investigator afloat. Since, following Belcher’s orders, Kellett had abandoned HMS Resolute and Intrepid, rather than their return journey ending near Cape Cockburn, Krabbé and his men had to continue for a further 140 nautical miles (260 km) to Beechey Island. This made the total length of their sledge trip 863½ nautical miles (1589 km), one of the longest man- hauled sledge trips in the history of the Arctic. Introduction On 22 July 2010 a party from the underwater archaeology division of Parks Canada flew into Mercy Bay in Aulavik National Park, on Banks Island, Northwest Territories – its mission to try to locate HMS Investigator, abandoned here by Commander Robert McClure in 1853.3 Two days later underwater archaeologists Ryan Harris and Jonathan Moore took to the water in a Zodiac to search the bay, towing a side-scan sonar towfish. -

THE SUN, the MOON and FIRMAMENT in CHUKCHI MYTHOLOGY and on the RELATIONS of CELESTIAL BODIES and SACRIFICE Ülo Siimets
THE SUN, THE MOON AND FIRMAMENT IN CHUKCHI MYTHOLOGY AND ON THE RELATIONS OF CELESTIAL BODIES AND SACRIFICE Ülo Siimets Abstract This article gives a brief overview of the most common Chukchi myths, notions and beliefs related to celestial bodies at the end of the 19th and during the 20th century. The firmament of Chukchi world view is connected with their main source of subsistence – reindeer herding. Chukchis are one of the very few Siberian indigenous people who have preserved their religion. Similarly to many other nations, the peoples of the Far North as well as Chukchis personify the Sun, the Moon and stars. The article also points out the similarities between Chukchi notions and these of other peoples. Till now Chukchi reindeer herders seek the supposed help or influence of a constellation or planet when making important sacrifices (for example, offering sacrifices in a full moon). According to the Chukchi religion the most important celestial character is the Sun. It is spoken of as an individual being (vaúrgún). In addition to the Sun, the Creator, Dawn, Zenith, Midday and the North Star also belong to the ranks of special (superior) beings. The Moon in Chukchi mythology is a man and a being in one person. It is as the ketlja (evil spirit) of the Sun. Chukchi myths about several stars (such as the North Star and Betelgeuse) resemble to a great extent these of other peoples. Keywords: astral mythology, the Moon, sacrifices, reindeer herding, the Sun, celestial bodies, Chukchi religion, constellations. The interdependence of the Earth and celestial as well as weather phenomena has a special meaning for mankind for it is the co-exist- ence of the Sun and Moon, day and night, wind, rainfall and soil that creates life and warmth and provides the daily bread. -

Rather Than Imposing Thematic Unity Or Predefining a Common Theoretical
The Supernatural Arctic: An Exploration Shane McCorristine, University College Dublin Abstract The magnetic attraction of the North exposed a matrix of motivations for discovery service in nineteenth-century culture: dreams of wealth, escape, extreme tourism, geopolitics, scientific advancement, and ideological attainment were all prominent factors in the outfitting expeditions. Yet beneath this „exoteric‟ matrix lay a complex „esoteric‟ matrix of motivations which included the compelling themes of the sublime, the supernatural, and the spiritual. This essay, which pivots around the Franklin expedition of 1845-1848, is intended to be an exploration which suggests an intertextuality across Arctic time and geography that was co-ordinated by the lure of the supernatural. * * * Introduction In his classic account of Scott‟s Antarctic expedition Apsley Cherry- Garrard noted that “Polar exploration is at once the cleanest and most isolated way of having a bad time which has been devised”.1 If there is one single question that has been asked of generations upon generations of polar explorers it is, Why?: Why go through such ordeals, experience such hardship, and take such risks in order to get from one place on the map to another? From an historical point of view, with an apparent fifty per cent death rate on polar voyages in the long nineteenth century amid disaster after disaster, the weird attraction of the poles in the modern age remains a curious fact.2 It is a less curious fact that the question cui bono? also featured prominently in Western thinking about polar exploration, particularly when American expeditions entered the Arctic 1 Apsley Cherry-Garrard, The Worst Journey in the World. -

Meaningful Consultation, Meaningful Participants and Meaning Making: Inuvialuit Perspectives on the Mackenzie Valley Pipeline and the Climate Crisis
Meaningful Consultation, Meaningful Participants and Meaning Making: Inuvialuit Perspectives on the Mackenzie Valley Pipeline and the Climate Crisis by Letitia Pokiak BA Anthropology, from the University of Alberta, 2003 A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of MASTER OF ARTS in the Department of Anthropology © Letitia Pokiak, 2020 University of Victoria All rights reserved. This Thesis may not be reproduced in whole or in part, by photocopy or other means, without the permission of the author. We acknowledge with respect the Lekwungen peoples on whose traditional territory the university stands and the Songhees, Esquimalt and WSÁNEĆ peoples whose historical relationships with the land continue to this day. Supervisory Committee Meaningful Consultation, Meaningful Participants and Meaning Making: Inuvialuit Perspectives on the Mackenzie Valley Pipeline and the Climate Crisis by Letitia Pokiak BA ANTHROPOLOGY, from the University of Alberta, 2003 Supervisory Committee Dr. Brian Thom, Department of Anthropology Supervisor Dr. Robert Hancock, Department of Anthropology Departmental Member ii Abstract This Inuvialuit ‘story’ revolves around the Inuvialuit uprising and resurgence against government and industrial encroachment, and the self determination efforts to regain sovereignty of traditional territories. This ‘story’ also discusses how meaningful consultation made the Inuvialuit Final Agreement a reality, through which Inuvialuit land rights and freedoms were formally acknowledged and entrenched in the Canadian Constitution. Through meaningful consultation, Inuvialuit have become ‘meaningful participants’ in sustainable and future-making decisions of Inuvialuit nunangat (Inuvialuit lands) and waters, with respect to the Inuvialuit People and natural beings that Inuvialuit depend upon and maintain relationship with. As ‘meaningful participants’, Inuvialuit have the sovereign rights to “make meaning” and carve out a future as a sovereign nation within the country of Canada. -

About the Caa / À Propos De L'aca
ABOUT THE CAA / À PROPOS DE L’ACA The Canadian Archaeological Association (caa) was founded in 1968. Membership includes professional, avocational and student archaeologists, as well as individuals of the general public of any country, who are interested in furthering the objectives of the Association. The objectives of the caa are as follows: § To promote the increase and the dissemination of archaeological knowledge in Canada; § To promote active discourse and cooperation among archaeological societies and agencies and encourage archaeological research and conservation efforts; § To foster cooperative endeavours with aboriginal groups and agencies concerned with First Peoples’ heritage of Canada; § To serve as the national association capable of promoting activities advantageous to archaeology and discouraging activities detrimental to archaeology; § To publish archaeological literature, and; § To stimulate the interest of the general public in archaeology. ——— L’Association canadienne d’archéologie (aca) a été fondée en 1968. Ses adhérents comptent des archéologues dont c’est la profession ou un violon d’Ingres et des étudiants, ainsi que des membres venant du grand public et de n’importe quel pays, qui ont en vue de favoriser les objectifs de l’Association. Les objectifs de l’aca sont les suivants: § promouvoir l’accroissement et la propagation de connaissances archéologiques au Canada; § promouvoir une coopération et des échanges actifs entre les sociétés et les organismes archéologiques, et favoriser le travail de recherche et de conservation; § stimuler les efforts de coopération avec les groupes autochtones et les organismes concernés par le patrimoine canadien des Premières nations; § servir d’association nationale pouvant promouvoir les activités avantageuses pour l’archéologie et décourager les activités nuisibles à l’archéologie; § publier de la documentation archéologique; § stimuler l’intérêt du grand public pour l’archéologie.