AEBR 102 Review of Research and Monitoring Studies on New Zealand
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Notoraja Martinezi Sp. Nov., a New Species of Deepwater Skate and The
Zootaxa 4098 (1): 179–190 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2016 Magnolia Press ISSN 1175-5334 (online edition) http://doi.org/10.11646/zootaxa.4098.1.9 http://zoobank.org/urn:lsid:zoobank.org:pub:C7826AC7-6493-42D3-A0A4-89FEBB5AB3E3 Notoraja martinezi sp. nov., a new species of deepwater skate and the first record of the genus Notoraja Ishiyama, 1958 (Rajiformes: Arhynchobatidae) from the eastern Pacific Ocean FRANCISCO J. CONCHA1,2,7, DAVID A. EBERT3,4,5 & DOUGLAS J. LONG4,6 1Department of Ecology & Evolutionary Biology, University of Connecticut. 75 North Eagleville Road – Unit 3043 Storrs, CT, 06269, USA. E-mail: [email protected] 2Facultad de Ciencias del Mar y de Recursos Naturales, Universidad de Valparaíso. Av. Borgoño 16344, Viña del Mar, Chile. 3Pacific Shark Research Center, Moss Landing Marine Laboratories, Moss Landing, CA 95039, USA. 4Department of Ichthyology, California Academy of Sciences, 55 Music Concourse Drive, San Francisco, CA. 94118, USA. 5South African Institute for Aquatic Biodiversity, Private Bag 1015, Grahamstown, 6140, South Africa. E-mail: [email protected] state.edu. 6Department of Biology, St. Mary’s College, 1928 St. Mary’s Road, Moraga, California 94575. E-mail: [email protected] 7Corresponding author Abstract A new arhynchobatid skate, Notoraja martinezi, sp. nov., is described from four specimens collected from the eastern Central Pacific from Costa Rica to Ecuador and between depths of 1256–1472 m. The new species is placed in the genus Notoraja based on the long and flexible rostrum and its proportionally long tail with respect to total length. -

Bibliography Database of Living/Fossil Sharks, Rays and Chimaeras (Chondrichthyes: Elasmobranchii, Holocephali) Papers of the Year 2016
www.shark-references.com Version 13.01.2017 Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichthyes: Elasmobranchii, Holocephali) Papers of the year 2016 published by Jürgen Pollerspöck, Benediktinerring 34, 94569 Stephansposching, Germany and Nicolas Straube, Munich, Germany ISSN: 2195-6499 copyright by the authors 1 please inform us about missing papers: [email protected] www.shark-references.com Version 13.01.2017 Abstract: This paper contains a collection of 803 citations (no conference abstracts) on topics related to extant and extinct Chondrichthyes (sharks, rays, and chimaeras) as well as a list of Chondrichthyan species and hosted parasites newly described in 2016. The list is the result of regular queries in numerous journals, books and online publications. It provides a complete list of publication citations as well as a database report containing rearranged subsets of the list sorted by the keyword statistics, extant and extinct genera and species descriptions from the years 2000 to 2016, list of descriptions of extinct and extant species from 2016, parasitology, reproduction, distribution, diet, conservation, and taxonomy. The paper is intended to be consulted for information. In addition, we provide information on the geographic and depth distribution of newly described species, i.e. the type specimens from the year 1990- 2016 in a hot spot analysis. Please note that the content of this paper has been compiled to the best of our abilities based on current knowledge and practice, however, -

First Record of Swimming Speed of the Pacific Sleeper Shark Somniosus
Journal of the Marine First record of swimming speed of the Pacific Biological Association of the United Kingdom sleeper shark Somniosus pacificus using a baited camera array cambridge.org/mbi Yoshihiro Fujiwara , Yasuyuki Matsumoto, Takumi Sato, Masaru Kawato and Shinji Tsuchida Original Article Research Institute for Global Change (RIGC), Japan Agency for Marine-Earth Science and Technology (JAMSTEC), 2-15 Yokosuka, Kanagawa 237-0061, Japan Cite this article: Fujiwara Y, Matsumoto Y, Sato T, Kawato M, Tsuchida S (2021). First record of swimming speed of the Pacific Abstract sleeper shark Somniosus pacificus using a baited camera array. Journal of the Marine The Pacific sleeper shark Somniosus pacificus is one of the largest predators in deep Suruga Biological Association of the United Kingdom Bay, Japan. A single individual of the sleeper shark (female, ∼300 cm in total length) was 101, 457–464. https://doi.org/10.1017/ observed with two baited camera systems deployed simultaneously on the deep seafloor in S0025315421000321 the bay. The first arrival was recorded 43 min after the deployment of camera #1 on 21 July 2016 at a depth of 609 m. The shark had several remarkable features, including the Received: 26 July 2020 Revised: 14 April 2021 snout tangled in a broken fishing line, two torn anteriormost left-gill septums, and a parasitic Accepted: 14 April 2021 copepod attached to each eye. The same individual appeared at camera #2, which was First published online: 18 May 2021 deployed at a depth of 603 m, ∼37 min after it disappeared from camera #1 view. Finally, the same shark returned to camera #1 ∼31 min after leaving camera #2. -

Monitoring Elasmobranch Assemblages in a Data-Poor Country from the Eastern Tropical Pacific Using Baited Remote Underwater Vide
www.nature.com/scientificreports OPEN Monitoring elasmobranch assemblages in a data‑poor country from the Eastern Tropical Pacifc using baited remote underwater video stations Mario Espinoza1,2,3*, Tatiana Araya‑Arce1,2, Isaac Chaves‑Zamora1,2,4, Isaac Chinchilla5 & Marta Cambra1,2 Understanding how threatened species are distributed in space and time can have direct applications to conservation planning. However, implementing standardized methods to monitor populations of wide‑ranging species is often expensive and challenging. In this study, we used baited remote underwater video stations (BRUVS) to quantify elasmobranch abundance and distribution patterns across a gradient of protection in the Pacifc waters of Costa Rica. Our BRUVS survey detected 29 species, which represents 54% of the entire elasmobranch diversity reported to date in shallow waters (< 60 m) of the Pacifc of Costa Rica. Our data demonstrated that elasmobranchs beneft from no‑take MPAs, yet large predators are relatively uncommon or absent from open‑fshing sites. We showed that BRUVS are capable of providing fast and reliable estimates of the distribution and abundance of data‑poor elasmobranch species over large spatial and temporal scales, and in doing so, they can provide critical information for detecting population‑level changes in response to multiple threats such as overfshing, habitat degradation and climate change. Moreover, given that 66% of the species detected are threatened, a well‑designed BRUVS survey may provide crucial population data for assessing the conservation status of elasmobranchs. These eforts led to the establishment of a national monitoring program focused on elasmobranchs and key marine megafauna that could guide monitoring eforts at a regional scale. -

© Iccat, 2007
A5 By-catch Species APPENDIX 5: BY-CATCH SPECIES A.5 By-catch species By-catch is the unintentional/incidental capture of non-target species during fishing operations. Different types of fisheries have different types and levels of by-catch, depending on the gear used, the time, area and depth fished, etc. Article IV of the Convention states: "the Commission shall be responsible for the study of the population of tuna and tuna-like fishes (the Scombriformes with the exception of Trichiuridae and Gempylidae and the genus Scomber) and such other species of fishes exploited in tuna fishing in the Convention area as are not under investigation by another international fishery organization". The following is a list of by-catch species recorded as being ever caught by any major tuna fishery in the Atlantic/Mediterranean. Note that the lists are qualitative and are not indicative of quantity or mortality. Thus, the presence of a species in the lists does not imply that it is caught in significant quantities, or that individuals that are caught necessarily die. Skates and rays Scientific names Common name Code LL GILL PS BB HARP TRAP OTHER Dasyatis centroura Roughtail stingray RDC X Dasyatis violacea Pelagic stingray PLS X X X X Manta birostris Manta ray RMB X X X Mobula hypostoma RMH X Mobula lucasana X Mobula mobular Devil ray RMM X X X X X Myliobatis aquila Common eagle ray MYL X X Pteuromylaeus bovinus Bull ray MPO X X Raja fullonica Shagreen ray RJF X Raja straeleni Spotted skate RFL X Rhinoptera spp Cownose ray X Torpedo nobiliana Torpedo -

BETTER DATA COLLECTION in SHARK FISHERIES LEARNING from PRACTICE Cover Image: Cacaodesign.It FAO Fisheries and Aquaculture Circular No
NFIF/C1227 (En) FAO Fisheries and Aquaculture Circular ISSN 2070-6065 BETTER DATA COLLECTION IN SHARK FISHERIES LEARNING FROM PRACTICE Cover image: cacaodesign.it FAO Fisheries and Aquaculture Circular No. 1227 NFIF/C1227 (En) BETTER DATA COLLECTION IN SHARK FISHERIES LEARNING FROM PRACTICE by Monica Barone Fishery Resources Consultant FAO Fisheries Department Rome, Italy and Kim Friedman Senior Fishery Resources Officer FAO Fisheries Department Rome, Italy FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS Rome, 2021 Required citation: FAO. 2021. Better data collection in shark fisheries – Learning from practice. FAO Fisheries and Aquaculture Circular No. 1227. Rome. https://doi.org/10.4060/cb5378en The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. The views expressed in this information product are those of the author(s) and do not necessarily reflect the views or policies of FAO. ISBN 978-92-5-134622-8 © FAO, 2021 Some rights reserved. This work is made available under the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 IGO licence (CC BY-NC-SA 3.0 IGO; https://creativecommons.org/licenses/by-nc-sa/3.0/igo/legalcode). -

Characterization of the Artisanal Elasmobranch Fisheries Off The
3 National Marine Fisheries Service Fishery Bulletin First U.S. Commissioner established in 1881 of Fisheries and founder NOAA of Fishery Bulletin Abstract—The landings of the artis- Characterization of the artisanal elasmobranch anal elasmobranch fisheries of 3 com- munities located along the Pacific coast fisheries off the Pacific coast of Guatemala of Guatemala from May 2017 through March 2020 were evaluated. Twenty- Cristopher G. Avalos Castillo (contact author)1,2 one elasmobranch species were iden- 3,4 tified in this study. Bottom longlines Omar Santana Morales used for multispecific fishing captured ray species and represented 59% of Email address for contact author: [email protected] the fishing effort. Gill nets captured small shark species and represented 1 Fundación Mundo Azul 3 Facultad de Ciencias Marinas 41% of the fishing effort. The most fre- Carretera a Villa Canales Universidad Autónoma de Baja California quently caught species were the longtail km 21-22 Finca Moran Carretera Ensenada-Tijuana 3917 stingray (Hypanus longus), scalloped 01069 Villa Canales, Guatemala Fraccionamiento Playitas hammerhead (Sphyrna lewini), and 2 22860 Ensenada, Baja California, Mexico Pacific sharpnose shark (Rhizopriono- Centro de Estudios del Mar y Acuicultura 4 don longurio), accounting for 47.88%, Universidad de San Carlos de Guatemala ECOCIMATI A.C. 33.26%, and 7.97% of landings during Ciudad Universitaria Zona 12 Avenida del Puerto 2270 the monitoring period, respectively. Edificio M14 Colonia Hidalgo The landings were mainly neonates 01012 Guatemala City, Guatemala 22880 Ensenada, Baja California, Mexico and juveniles. Our findings indicate the presence of nursery areas on the continental shelf off Guatemala. -

New Zealand Fishes a Field Guide to Common Species Caught by Bottom, Midwater, and Surface Fishing Cover Photos: Top – Kingfish (Seriola Lalandi), Malcolm Francis
New Zealand fishes A field guide to common species caught by bottom, midwater, and surface fishing Cover photos: Top – Kingfish (Seriola lalandi), Malcolm Francis. Top left – Snapper (Chrysophrys auratus), Malcolm Francis. Centre – Catch of hoki (Macruronus novaezelandiae), Neil Bagley (NIWA). Bottom left – Jack mackerel (Trachurus sp.), Malcolm Francis. Bottom – Orange roughy (Hoplostethus atlanticus), NIWA. New Zealand fishes A field guide to common species caught by bottom, midwater, and surface fishing New Zealand Aquatic Environment and Biodiversity Report No: 208 Prepared for Fisheries New Zealand by P. J. McMillan M. P. Francis G. D. James L. J. Paul P. Marriott E. J. Mackay B. A. Wood D. W. Stevens L. H. Griggs S. J. Baird C. D. Roberts‡ A. L. Stewart‡ C. D. Struthers‡ J. E. Robbins NIWA, Private Bag 14901, Wellington 6241 ‡ Museum of New Zealand Te Papa Tongarewa, PO Box 467, Wellington, 6011Wellington ISSN 1176-9440 (print) ISSN 1179-6480 (online) ISBN 978-1-98-859425-5 (print) ISBN 978-1-98-859426-2 (online) 2019 Disclaimer While every effort was made to ensure the information in this publication is accurate, Fisheries New Zealand does not accept any responsibility or liability for error of fact, omission, interpretation or opinion that may be present, nor for the consequences of any decisions based on this information. Requests for further copies should be directed to: Publications Logistics Officer Ministry for Primary Industries PO Box 2526 WELLINGTON 6140 Email: [email protected] Telephone: 0800 00 83 33 Facsimile: 04-894 0300 This publication is also available on the Ministry for Primary Industries website at http://www.mpi.govt.nz/news-and-resources/publications/ A higher resolution (larger) PDF of this guide is also available by application to: [email protected] Citation: McMillan, P.J.; Francis, M.P.; James, G.D.; Paul, L.J.; Marriott, P.; Mackay, E.; Wood, B.A.; Stevens, D.W.; Griggs, L.H.; Baird, S.J.; Roberts, C.D.; Stewart, A.L.; Struthers, C.D.; Robbins, J.E. -
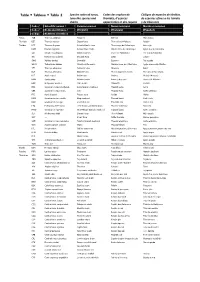
Table Tableau Tabla 2
Table Tableau Tabla 2 Species codes of tunas, Codes des espèces de Códigos de especies de túnidos, tuna‐like species and thonidés, d’espèces de especies afines a los túnidos sharks apparentées et des requins y de tiburones Code / Scientific names / Common names Noms communs Nombres comunes Code / Noms sientifiques / (English) (Français) (Español) Código Nombres científicos Tunas ALB Thunnus alalunga Albacore Germon Atún blanco Thonidés BET Thunnus obesus Bigeye tuna Thon obèse(=Patudo) Patudo Túnidos BFT Thunnus thynnus Atlantic bluefin tuna Thon rouge de l’atlantique Atún rojo BUM Makaira nigricans Atlantic blue marlin Makaire bleu de l'Atlantique Aguja azul del Atlántico SAI Istiophorus albicans Atlantic sailfish Voilier de l'Atlantique Pez vela del Atlántico SKJ Katsuwonus pelamis Skipjack tuna Listao Listado SWO Xiphias gladius Swordfish Espadon Pez espada WHM Tetrapturus albidus Atlantic white marlin Makaire blanc de l'Atlantique Aguja blanca del Atlántico YFT Thunnus albacares Yellowfin tuna Albacore Rabil BLF Thunnus atlanticus Blackfin tuna Thon à nageoires noires Atún des aletas negras BLT Auxis rochei Bullet tuna Bonitou Melva(=Melvera) BON Sarda sarda Atlantic bonito Bonite à dos rayé Bonito del Atlántico BOP Orcynopsis unicolor Plain bonito Palomette Tasarte BRS Scomberomorus brasiliensis Serra Spanish mackerel Thazard serra Serra CER Scomberomorus regalis Cero Thazard franc Carite chinigua FRI Auxis thazard Frigate tuna Auxide Melva KGM Scomberomorus cavalla King mackerel Thazard barré Carite lucio KGX Scomberomorus spp -

Wk Shark Advice Adhshark
ICES Special Request Advice Ecoregions in the Northeast Atlantic and adjacent seas Published 25 September 2020 NEAFC and OSPAR joint request on the status and distribution of deep-water elasmobranchs Advice summary In response to a joint request from NEAFC and OSPAR, ICES reviewed existing information on deep-water sharks, skates and rays from surveys and the literature. Distribution maps were generated for 21 species, showing the location of catches from available survey data on deep-water sharks and elasmobranchs in the NEAFC and OSPAR areas of the Northeast Atlantic. Shapefiles of the species distribution areas are available as supporting documentation to this work. This advice sheet presents a summary of ICES advice on the stock status of species for which an assessment is available, as well as current knowledge on the stock status of species for which ICES does not provide advice. An overview of approaches which may be applied to mitigate bycatch and to improve stock status is also presented. ICES recognizes that, despite their limitations, prohibition, gear and depth limitations, and TAC are mechanisms currently available to managers to regulate outtake; therefore, ICES advises that these mechanisms should be maintained. Furthermore, ICES advises that additional measures, such as electromagnetic exclusion devices, acoustic or light-based deterrents, and spatio-temporal management could be explored. Request NEAFC and OSPAR requested ICES to produce: a. Maps and shapefiles of the distribution of the species, identifying, if possible, key areas used during particular periods/stages of the species’ lifecycle in terms of distribution and relative abundance of the species, and expert interpretation of the data products; b. -

2018 Final LOFF W/ Ref and Detailed Info
Final List of Foreign Fisheries Rationale for Classification ** (Presence of mortality or injury (P/A), Co- Occurrence (C/O), Company (if Source of Marine Mammal Analogous Gear Fishery/Gear Number of aquaculture or Product (for Interactions (by group Marine Mammal (A/G), No RFMO or Legal Target Species or Product Type Vessels processor) processing) Area of Operation or species) Bycatch Estimates Information (N/I)) Protection Measures References Detailed Information Antigua and Barbuda Exempt Fisheries http://www.fao.org/fi/oldsite/FCP/en/ATG/body.htm http://www.fao.org/docrep/006/y5402e/y5402e06.htm,ht tp://www.tradeboss.com/default.cgi/action/viewcompan lobster, rock, spiny, demersal fish ies/searchterm/spiny+lobster/searchtermcondition/1/ , (snappers, groupers, grunts, ftp://ftp.fao.org/fi/DOCUMENT/IPOAS/national/Antigua U.S. LoF Caribbean spiny lobster trap/ pot >197 None documented, surgeonfish), flounder pots, traps 74 Lewis Fishing not applicable Antigua & Barbuda EEZ none documented none documented A/G AndBarbuda/NPOA_IUU.pdf Caribbean mixed species trap/pot are category III http://www.nmfs.noaa.gov/pr/interactions/fisheries/tabl lobster, rock, spiny free diving, loops 19 Lewis Fishing not applicable Antigua & Barbuda EEZ none documented none documented A/G e2/Atlantic_GOM_Caribbean_shellfish.html Queen conch (Strombus gigas), Dive (SCUBA & free molluscs diving) 25 not applicable not applicable Antigua & Barbuda EEZ none documented none documented A/G U.S. trade data Southeastern U.S. Atlantic, Gulf of Mexico, and Caribbean snapper- handline, hook and grouper and other reef fish bottom longline/hook-and-line/ >5,000 snapper line 71 Lewis Fishing not applicable Antigua & Barbuda EEZ none documented none documented N/I, A/G U.S. -

Downloaded from the CCAMLR Website at the CCAMLR Scheme of International Scientific Observation Webpage (
Scheme of International Scientific Observation Scientific Observer’s Manual Finfish Fisheries Version 2020 Address 181 Macquarie Street, Hobart, Tasmania 7000, Australia Web www.ccamlr.org Telephone +61 3 6210 1110 Post PO Box 213, North Hobart, Tasmania 7002, Australia Email [email protected] Fax +61 3 6224 8744 This manual is produced in the official languages of the Commission (English, French, Russian and Spanish) and may be downloaded from the CCAMLR website at the CCAMLR Scheme of International Scientific Observation webpage (www.ccamlr.org/node/73033). Version Release date Observer forms covered Description 2011 01/12/2010 2011 – 2015 Longline Original 2011 – 2015 Finfish Trawl 2011 – 2015 Krill Trawl 2019 Draft 01/10/2018 2019 Longline Draft version presented at 2019 Finfish Trawl WG-FSA-18 for review by 2019 Krill Trawl Members 2020 For release: 2020 Longline Presented to WG-EMM-2019 for 01/09/2019 2019 Finfish Trawl endorsement 2 Table of Contents 1. Introduction 5 2. SISO observer roles and responsibilities 5 3. Definition of terms 6 4. CCAMLR regulations 7 5. General operational procedures 8 6. Units and formats 9 7. Standard measurements 9 7.1 Fish 9 7.2 Skates and rays 10 8. Weights 11 9. Sexing and maturity stages 11 9.1 Toothfish 11 9.2 Skates and rays 14 10. Otolith collection and storage 15 10.1 Removing otoliths 15 10.2 Otolith storage 18 11. Conversion factor tests – finfish fisheries 18 12. Target Species Identification Guide 19 12.1 Toothfish 19 12.2 Patagonian toothfish 19 12.3 Antarctic Toothfish 20 12.4 Toothfish species differences 20 12.5 Mackerel icefish 22 13.