Estimating Collateral Mortality from Towed Fishing Gear
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Educators' Resource Guide
EDUCATORS' RESOURCE GUIDE Produced and published by 3D Entertainment Distribution Written by Dr. Elisabeth Mantello In collaboration with Jean-Michel Cousteau’s Ocean Futures Society TABLE OF CONTENTS TO EDUCATORS .................................................................................................p 3 III. PART 3. ACTIVITIES FOR STUDENTS INTRODUCTION .................................................................................................p 4 ACTIVITY 1. DO YOU Know ME? ................................................................. p 20 PLANKton, SOURCE OF LIFE .....................................................................p 4 ACTIVITY 2. discoVER THE ANIMALS OF "SECRET OCEAN" ......... p 21-24 ACTIVITY 3. A. SECRET OCEAN word FIND ......................................... p 25 PART 1. SCENES FROM "SECRET OCEAN" ACTIVITY 3. B. ADD color to THE octoPUS! .................................... p 25 1. CHristmas TREE WORMS .........................................................................p 5 ACTIVITY 4. A. WHERE IS MY MOUTH? ..................................................... p 26 2. GIANT BasKET Star ..................................................................................p 6 ACTIVITY 4. B. WHat DO I USE to eat? .................................................. p 26 3. SEA ANEMONE AND Clown FISH ......................................................p 6 ACTIVITY 5. A. WHO eats WHat? .............................................................. p 27 4. GIANT CLAM AND ZOOXANTHELLAE ................................................p -

Sturgeon Chub (Macrhybopsis Gelida): a Technical Conservation Assessment
Sturgeon Chub (Macrhybopsis gelida): A Technical Conservation Assessment Prepared for the USDA Forest Service, Rocky Mountain Region, Species Conservation Project August 31, 2004 Frank J. Rahel and Laura A. Thel Department of Zoology and Physiology University of Wyoming, Laramie, Wyoming 82071 Peer Review Administered by American Fisheries Society Rahel, F.J. and L.A. Thel. (2004, August 31). Sturgeon Chub (Macrhybopsis gelida): a technical conservation assessment. [Online]. USDA Forest Service, Rocky Mountain Region. Available: http://www.fs.fed.us/r2/ projects/scp/assessments/sturgeonchub.pdf [date of access]. ACKNOWLEDGEMENTS We thank biologists from Colorado, Kansas, Nebraska, South Dakota, and Wyoming, and from the national forests and national grasslands within Region 2 who provided information about sturgeon chub within their jurisdictions. We especially thank Gregory Hayward and Richard Vacirca of the USDA Forest Service for their review of this species assessment. Comments also were provided by two anonymous reviewers. David B. McDonald of the University of Wyoming provided the population demographic matrix analysis. AUTHORS’ BIOGRAPHIES Frank J. Rahel is a professor in the Department of Zoology and Physiology at the University of Wyoming where he teaches courses in fi sheries management, ichthyology, and conservation biology. His research interests are centered around fi sh ecology and the infl uence of anthropogenic disturbances on fi sh assemblages. Laura A. Thel is a graduate research assistant in the Department of Zoology and Physiology at the University of Wyoming with research interests involving stream ecology, hydrology, and landscape ecology, especially as these are related to the management of native fi shes. COVER PHOTO CREDIT Sturgeon Chub (Macrhybopsis gelida). -

Petition to List the Black Teatfish, Holothuria Nobilis, Under the U.S. Endangered Species Act
Before the Secretary of Commerce Petition to List the Black Teatfish, Holothuria nobilis, under the U.S. Endangered Species Act Photo Credit: © Philippe Bourjon (with permission) Center for Biological Diversity 14 May 2020 Notice of Petition Wilbur Ross, Secretary of Commerce U.S. Department of Commerce 1401 Constitution Ave. NW Washington, D.C. 20230 Email: [email protected], [email protected] Dr. Neil Jacobs, Acting Under Secretary of Commerce for Oceans and Atmosphere U.S. Department of Commerce 1401 Constitution Ave. NW Washington, D.C. 20230 Email: [email protected] Petitioner: Kristin Carden, Oceans Program Scientist Sarah Uhlemann, Senior Att’y & Int’l Program Director Center for Biological Diversity Center for Biological Diversity 1212 Broadway #800 2400 NW 80th Street, #146 Oakland, CA 94612 Seattle,WA98117 Phone: (510) 844‐7100 x327 Phone: (206) 324‐2344 Email: [email protected] Email: [email protected] The Center for Biological Diversity (Center, Petitioner) submits to the Secretary of Commerce and the National Oceanographic and Atmospheric Administration (NOAA) through the National Marine Fisheries Service (NMFS) a petition to list the black teatfish, Holothuria nobilis, as threatened or endangered under the U.S. Endangered Species Act (ESA), 16 U.S.C. § 1531 et seq. Alternatively, the Service should list the black teatfish as threatened or endangered throughout a significant portion of its range. This species is found exclusively in foreign waters, thus 30‐days’ notice to affected U.S. states and/or territories was not required. The Center is a non‐profit, public interest environmental organization dedicated to the protection of native species and their habitats. -

IGFA Angling Rules
International Angling Rules The following angling rules have been formulated by the International Game Fish Association to promote ethical and sporting angling practices, to establish uniform regulations for the compilation of world game fish records, and to provide basic angling guidelines for use in fishing tournaments and any other group angling activities. The word “angling” is defined as catching or attempting to catch fish with a rod, reel, line, and hook as outlined in the international angling rules. There are some aspects of angling that cannot be controlled through rule making, however. Angling regulations cannot insure an outstanding performance from each fish, and world records cannot indicate the amount of difficulty in catching the fish. Captures in which the fish has not fought or has not had a chance to fight do not reflect credit on the fisherman, and only the angler can properly evaluate the degree of achievement in establishing the record. Only fish caught in accordance with IGFA international angling rules, and within the intent of these rules, will be considered for world records. Following are the rules for freshwater and saltwater fishing and a separate set of rules for fly fishing. RULES FOR FISHING IN FRESH AND SALT WATER (Also see Rules for Fly fishing) E. ROD Equipment Regulations 1. Rods must comply with sporting ethics and customs. A. LINE Considerable latitude is allowed in the choice of a rod, but rods giving 1. Monofilament, multifilament, and lead core multifilament the angler an unfair advantage will be disqualified. This rule is lines may be used. For line classes, see World Record Requirements. -
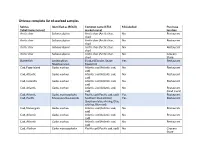
Ottawa: Complete List of Seafood Samples
Ottawa: complete list of seafood samples Sold as Identified as (BOLD) Common name (CFIA Mislabelled Purchase (label/menu/server) market name) location Arctic char Salverus alpirus Arctic char (Arctic char, No Restaurant char) Arctic char Salverus alpirus Arctic char (Arctic char, No Restaurant char) Arctic char Salverus alpirus Arctic char (Arctic char, No Restaurant char) Arctic char Salverus alpirus Arctic char (Arctic char, No Grocery char) Store Butterfish Lepidocybium Escolar (Escolar, Snake Yes Restaurant flavobrunneum Mackerel) Cod, Fogo Island Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Icelandic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) (food truck) Cod, Atlantic Gadus macrocephalus Pacific cod (Pacific cod, cod) Yes Restaurant Cod, Pacific Micromesistius australis Southern blue whiting Yes Restaurant (Southern blue whiting, Blue whiting, Blue cod) Cod, Norwegian Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Alaskan Gadus macrocephalus Pacific cod (Pacific cod, cod) No Grocery Store Cod, Pacific Gadus macrocephalus Pacific cod (Pacific cod, cod) No Grocery Store Cod, North Atlantic Gadus macrocephalus Pacific cod (Pacific cod, cod) Yes Restaurant Cod Gadus macrocephalus Pacific cod (Pacific cod, cod) No Grocery Store Cod, Icelandic Gadus morhua Atlantic cod (Atlantic cod, No Grocery cod) Store Cod, Icelandic Gadus morhua Atlantic cod (Atlantic cod, No Grocery cod) Store Euro Bass Gadus morhua Atlantic cod (Atlantic cod, Yes Restaurant cod) Grouper Epinephelus diacanthus Spinycheek grouper (n/a) Yes – E. -

Marine Fish Conservation Global Evidence for the Effects of Selected Interventions
Marine Fish Conservation Global evidence for the effects of selected interventions Natasha Taylor, Leo J. Clarke, Khatija Alliji, Chris Barrett, Rosslyn McIntyre, Rebecca0 K. Smith & William J. Sutherland CONSERVATION EVIDENCE SERIES SYNOPSES Marine Fish Conservation Global evidence for the effects of selected interventions Natasha Taylor, Leo J. Clarke, Khatija Alliji, Chris Barrett, Rosslyn McIntyre, Rebecca K. Smith and William J. Sutherland Conservation Evidence Series Synopses 1 Copyright © 2021 William J. Sutherland This work is licensed under a Creative Commons Attribution 4.0 International license (CC BY 4.0). This license allows you to share, copy, distribute and transmit the work; to adapt the work and to make commercial use of the work providing attribution is made to the authors (but not in any way that suggests that they endorse you or your use of the work). Attribution should include the following information: Taylor, N., Clarke, L.J., Alliji, K., Barrett, C., McIntyre, R., Smith, R.K., and Sutherland, W.J. (2021) Marine Fish Conservation: Global Evidence for the Effects of Selected Interventions. Synopses of Conservation Evidence Series. University of Cambridge, Cambridge, UK. Further details about CC BY licenses are available at https://creativecommons.org/licenses/by/4.0/ Cover image: Circling fish in the waters of the Halmahera Sea (Pacific Ocean) off the Raja Ampat Islands, Indonesia, by Leslie Burkhalter. Digital material and resources associated with this synopsis are available at https://www.conservationevidence.com/ -

Wgbeam Report 2013
ICES WGBEAM REPORT 2013 SCICOM STEERING GROUP ON ECOSYSTEM SURVEYS SCIENCE AND TECHNOLOGY ICES CM 2013/SSGESST:12 REF. SCICOM & ACOM Report of the Working Group on Beam Trawl Surveys (WGBEAM) 23-26 April 2013 Ancona, Italy International Council for the Exploration of the Sea Conseil International pour l’Exploration de la Mer H. C. Andersens Boulevard 44–46 DK-1553 Copenhagen V Denmark Telephone (+45) 33 38 67 00 Telefax (+45) 33 93 42 15 www.ices.dk [email protected] Recommended format for purposes of citation: ICES. 2013. Report of the Working Group on Beam Trawl Surveys (WGBEAM), 23-26 April 2013, Ancona, Italy. ICES CM 2013/SSGESST:12. 260 pp. For permission to reproduce material from this publication, please apply to the Gen- eral Secretary. The document is a report of an Expert Group under the auspices of the International Council for the Exploration of the Sea and does not necessarily represent the views of the Council. © 2013 International Council for the Exploration of the Sea ICES WGBEAM REPORT 2013 | i Contents Executive summary ................................................................................................................ 1 1 Opening of the meeting ................................................................................................ 3 2 Adoption of the agenda ................................................................................................ 3 3 Introduction .................................................................................................................... 3 3.1 Terms of -

Traditional Ecological Knowledge in Andavadoaka, Southwest Madagascar
BLUE VENTURES CONSERVATION REPORT Josephine M. Langley Vezo Knowledge: Traditional Ecological Knowledge in Andavadoaka, southwest Madagascar 52 Avenue Road, London N6 5DR [email protected] Tel: +44 (0) 208 341 9819 Fax: +44 (0) 208 341 4821 BLUE VENTURES CONSERVATION REPORT Andavadoaka, 2006 © Blue Ventures 2006 Copyright in this publication and in all text, data and images contained herein, except as otherwise indicated, rests with Blue Ventures. Recommended citation: Langley, J. (2006). Vezo Knowledge: Traditional Ecological Knowledge in Andavadoaka, southwest Madagascar. BLUE VENTURES CONSERVATION REPORT Summary Many fisheries and marine conservation management (ii) Knowledge of marine resources is passed orally from projects throughout the world have been dogged by generation to generation failure as a result of a lack of acceptance of (iii) Traditional laws, taboos and ceremonies are commonly management interventions by local communities. used in natural resource management Evaluation of these failed studies has produced (iv) Lifestyle, food security and housing are all dependent extensive guidelines, manuals and new fields of study on natural resources and the use of coastal and marine (Bunce and Pomeroy, 2004). Community engagement, resources form an essential part of this participatory research, and promoting the use of local (v) The arrival of the Catholic Mission has reduced the knowledge have repeatedly emerged as steps necessary proportion of villagers adhering to traditional ancestor to address the problem of managing the development of worship people and their economies while simultaneously (vi) There has been a change in recent years from a barter protecting the environment (Berkes et al, 2001; Bunce and subsistence economy to a market-driven cash- based economy and Pomeroy, 2004; Wibera et al, 2004; Scholz et al, (vii) Increased income for some members of the 2004). -

Marine Biodiversity of the South East NRM Region
Marine Environment and Ecology Benthic Ecology Subprogram Marine Biodiversity of the South East NRM Region SARDI Publication No. F2009/000681-1 SARDI Research Report series No. 416 Keith Rowling, Shirley Sorokin, Leonardo Mantilla and David Currie SARDI Aquatic Sciences PO BOX 120 Henley Beach SA 5022 December 2009 Prepared for the Department for Environment and Heritage 1 Information Systems and Database Support Program Marine Biodiversity of the South East NRM Region Keith Rowling, Shirley Sorokin, Leonardo Mantilla and David Currie December 2009 SARDI Publication No. F2009/000681-1 SARDI Research Report Series No. 416 Prepared for the Department for Environment and Heritage 2 This Publication may be cited as: Rowling, K.P., Sorokin, S.J., Mantilla, L. & Currie, D.R.. (2009) Marine Biodiversity of the South East NRM Region. South Australian Research and Development Institute (Aquatic Sciences), Adelaide. SARDI Publication No. F2009/000681-1. South Australian Research and Development Institute SARDI Aquatic Sciences 2 Hamra Avenue West Beach SA 5024 Telephone: (08) 8207 5400 Facsimile: (08) 8207 5406 http://www.sardi.sa.gov.au DISCLAIMER The authors warrant that they have taken all reasonable care in producing this report. The report has been through the SARDI internal review process, and has been formally approved for release by the Chief of Division. Although all reasonable efforts have been made to ensure quality, SARDI does not warrant that the information in this report is free from errors or omissions. SARDI does not accept any liability for the contents of this report or for any consequences arising from its use or any reliance placed upon it. -

Fishes of Terengganu East Coast of Malay Peninsula, Malaysia Ii Iii
i Fishes of Terengganu East coast of Malay Peninsula, Malaysia ii iii Edited by Mizuki Matsunuma, Hiroyuki Motomura, Keiichi Matsuura, Noor Azhar M. Shazili and Mohd Azmi Ambak Photographed by Masatoshi Meguro and Mizuki Matsunuma iv Copy Right © 2011 by the National Museum of Nature and Science, Universiti Malaysia Terengganu and Kagoshima University Museum All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means without prior written permission from the publisher. Copyrights of the specimen photographs are held by the Kagoshima Uni- versity Museum. For bibliographic purposes this book should be cited as follows: Matsunuma, M., H. Motomura, K. Matsuura, N. A. M. Shazili and M. A. Ambak (eds.). 2011 (Nov.). Fishes of Terengganu – east coast of Malay Peninsula, Malaysia. National Museum of Nature and Science, Universiti Malaysia Terengganu and Kagoshima University Museum, ix + 251 pages. ISBN 978-4-87803-036-9 Corresponding editor: Hiroyuki Motomura (e-mail: [email protected]) v Preface Tropical seas in Southeast Asian countries are well known for their rich fish diversity found in various environments such as beautiful coral reefs, mud flats, sandy beaches, mangroves, and estuaries around river mouths. The South China Sea is a major water body containing a large and diverse fish fauna. However, many areas of the South China Sea, particularly in Malaysia and Vietnam, have been poorly studied in terms of fish taxonomy and diversity. Local fish scientists and students have frequently faced difficulty when try- ing to identify fishes in their home countries. During the International Training Program of the Japan Society for Promotion of Science (ITP of JSPS), two graduate students of Kagoshima University, Mr. -

Appendix 13.2 Marine Ecology and Biodiversity Baseline Conditions
THE LONDON RESORT PRELIMINARY ENVIRONMENTAL INFORMATION REPORT Appendix 13.2 Marine Ecology and Biodiversity Baseline Conditions WATER QUALITY 13.2.1. The principal water quality data sources that have been used to inform this study are: • Environment Agency (EA) WFD classification status and reporting (e.g. EA 2015); and • EA long-term water quality monitoring data for the tidal Thames. Environment Agency WFD Classification Status 13.2.2. The tidal River Thames is divided into three transitional water bodies as part of the Thames River Basin Management Plan (EA 2015) (Thames Upper [ID GB530603911403], Thames Middle [ID GB53060391140] and Thames Lower [ID GB530603911401]. Each of these waterbodies are classified as heavily modified waterbodies (HMWBs). The most recent EA assessment carried out in 2016, confirms that all three of these water bodies are classified as being at Moderate ecological potential (EA 2018). 13.2.3. The Thames Estuary at the London Resort Project Site is located within the Thames Middle Transitional water body, which is a heavily modified water body on account of the following designated uses (Cycle 2 2015-2021): • Coastal protection; • Flood protection; and • Navigation. 13.2.4. The downstream extent of the Thames Middle transitional water body is located approximately 12 km downstream of the Kent Project Site and 8 km downstream of the Essex Project Site near Lower Hope Point. Downstream of this location is the Thames Lower water body which extends to the outer Thames Estuary. 13.2.5. A summary of the current Thames Middle water body WFD status is presented in Table A13.2.1, together with those supporting elements that do not currently meet at least Good status and their associated objectives. -

The Mysid-Feeding Guild of Demersal Fishes in the Brackish Zone of the Westerschelde Estuary
Chapter 7 The Mysid feeding guild offishes in the Westerschelde 123 CHAPTER 7 THE MYSID-FEEDING GUILD OF DEMERSAL FISHES IN THE BRACKISH ZONE OF THE WESTERSCHELDE ESTUARY 63962 K. Hostens - J. Mees University of Gent, Depa rtment of Biology, Marine Biology Section, Ledeganckstraat 35, B-9000 Gent, Belgium Key words: stomach content, consumption, Mysidacea, flatfish, gobies, gadoids, clupeoids Published in Journal of Fish Biology 1999, 55: 704-719 Abstract. The demersal fish fauna of the mesohaline zone of the Westerschelde estuary (south -west Netherlands) was sampled intensively in the period 1990-1992. Almost 500 beam trawl samples were taken in both subtidal (330 samples) and intertidal (144 samples) habitats. These yielded 44 fish species, mostly as juveniles. The area was found to function as a nursery for several demersal fish species. and harboured large populations ofhyper - benthic mysids. Three gobies, three flatfish, one clupeoid and one gadoid dominated the fish fauna, while three mysid species were important components of the holohyperbenthos. From c. 1500 stomach contents of 25 fish species, 44 prey species were identified, the most abundant of which were also common in the hyperbenthal. The demersal fish community consisted of a group that foraged subtidally on fast-moving epi- and hyperbenthic prey (for example gadoids, gobies and clupeoids) and a group that foraged on slow-moving or sessile endobenthic organisms, mainly in intertidal areas (for example most flatfish species). Mysidacea occurred in >50 % stom - achs analysed and were taken as prey by 19 of the 25 fish species. Mysids were most important in the diets of Pomatoschistus minutus, P.